Пузыреплодник как формировать: Как обрезать пузыреплодник, пошаговая инструкция от А до Я
как правильно формировать крону куста? в 2022 году на ГудГрунт
Содержание
- Санитарная обрезка
- Декоративная стрижка спиреи
- Омоложение кустарника
- Использование спиреи в ландшафтном дизайне
Существует много способов создать необычную декоративную композицию из спиреи (так иначе называют пузыреплодник): обрезка с приданием кроне разнообразных форм, сочетание сортов с разной расцветкой и конфигурацией листьев. Декоративный вид кустарник имеет весь сезон: сначала участок украсят компактные кустики с листвой разнообразных оттенков, затем грозди цветов белой или розовой окраски, а осенью на их месте образуются плоды оригинальной формы. Ландшафтные дизайнеры по достоинству оценили растение – используя только пузыреплодник разных видов, они создают интересные и оригинальные композиции.
Санитарная обрезка
Пузыреплодник редко болеет и повреждается насекомыми, поэтому санитарная обрезка не составит большой сложности. Ранней весной удалите все сломанные или засохшие ветки, приглядитесь – вдруг какой-то кустик подхватил инфекцию или его атаковали вредители. После этого посмотрите, нет ли побегов, растущих внутрь куста. Такие ветки тоже следует срезать, они делают куст слишком густым – ветер не пройдет сквозь листву, и воздух будет застаиваться. На этом можете закончить работу.
Ранней весной удалите все сломанные или засохшие ветки, приглядитесь – вдруг какой-то кустик подхватил инфекцию или его атаковали вредители. После этого посмотрите, нет ли побегов, растущих внутрь куста. Такие ветки тоже следует срезать, они делают куст слишком густым – ветер не пройдет сквозь листву, и воздух будет застаиваться. На этом можете закончить работу.
Осенью после листопада проверьте, нет ли поврежденных фрагментов, уберите слабые неразвитые побеги и все ветки, которые вызывают у вас подозрение. Зимние холода перенесут только сильные здоровые отростки, все остальные ослабят пузыреплодник. При каждой стрижке, а особенно осенью, сожгите все удаленные участки. Если их оставить до весны, вредители обязательно устроят себе зимовку на сухой древесине.
Утеплять спирею после осенней обрезки не нужно. Укрывают только молодые и ослабленные экземпляры.
Иногда санитарную обрезку приходится проводить и летом. Увидев поврежденный побег, сразу удалите его, чтобы не портить декоративный вид растения и не создавать условий для развития инфекции.
Спирея украшает участок красивыми листьями разных оттенков. Иногда гибридное растение может выпустить ветку, покрытую зеленой листвой. Не надейтесь, что с возрастом молодые листочки изменят окраску и не будут отличаться от остальной кроны. Такие фрагменты нужно удалять: расцветку они уже не поменяют, а вид куста испортят.
Декоративная стрижка спиреи
Некоторые декоративные деревца и кустарники не нуждаются в обязательном формировании кроны – а вот пузыреплодник нужно стричь, даже если вы не любите четкие контуры и предпочитаете естественную красоту растений. В отличие от туи и других хвойных пород, которые в любом случае отрастят множество побегов и образуют плотную крону, спирея сама не даст такой красоты. Она выпустит несколько веток, сильно вытягивающихся в длину, и посчитает свою миссию завершенной.
Перед началом работы учтите, что придется резать ветки разной толщины, поэтому соберите и остро наточите все необходимые инструменты:
- нож;
- садовые ножницы;
- секатор;
- ножовку.
Чтобы стимулировать рост новых побегов, старым побегам нельзя давать сильно вытягиваться. После окончания цветения укоротите у молодых кустов ветки на 1/3. На второй год и все следующие сезоны побеги нужно обрезать уже на 2/3 их длины. Куст примет компактную ровную форму, а контур зависит от ваших целей. Растение хорошо переносит стрижку и сразу же начнет выпускать новые отростки. Крона станет гуще, но ветки растут настолько активно, что иногда корректировать форму приходится каждый месяц. Иногда хозяева старательно срезают старые побеги, выполняют все правила при формировании кроны, а куст остается рыхлым, дает очень мало новых отростков. Виновата в этом не неумелая стрижка, а плохая почва или ненадлежащий уход.
 Растение неприхотливо, но ему тоже нужна и влага, и питательные вещества. Устраните свои ошибки, иначе никакая обрезка не придаст пузыреплоднику декоративный вид.
Растение неприхотливо, но ему тоже нужна и влага, и питательные вещества. Устраните свои ошибки, иначе никакая обрезка не придаст пузыреплоднику декоративный вид.Совет
Если при санитарной или декоративной обрезке приходится удалять молодые сильные побеги, можете использовать их для размножения куста. Заготовьте черенки с 2-3 междоузлиями, срежьте все листья, чтобы не было интенсивного испарения влаги, и высадите в песок. Когда увидите набухшие молодые почки, посадите веточки на постоянное место. Больше всего им понравится расти рядом с материнским пузыреплодником, с которого были взяты отростки.
Омоложение кустарника
Старые кусты теряют декоративность. Если замечаете, что листва стала не такой густой и свежей, куст цветет скудно, а соцветия стали меньше, значит, пузыреплоднику требуется омолаживающая обрезка. Обычно эта процедура проводится через 5-6 лет после посадки. Заранее запаситесь садовым варом и пилой, поскольку удалять придется толстые стволы.
Чтобы стимулировать развитие новых побегов, нужно провести омолаживающую обрезку. Лучше выполнять эту работу весной, пока не начали распускаться почки, или осенью, когда опадут листья. Побеги, которые от возраста уже не могут выпускать густую листву и пышные соцветия, нужно спилить до основания. Все остальные стволы срезаются до ответвления первого побега.
Масштабная обрезка ослабляет куст – обеспечьте ему хороший уход, своевременные поливы и подкормки. Если процедура проводилась осенью, утеплите пузыреплодник соломой или укрывным материалом. В регионах с морозными малоснежными зимами лучше защитить от холодов и спирею, которую омолодили весной.
Использование спиреи в ландшафтном дизайне
Побеги кустарника растут очень быстро, после стрижки растение за один сезон станет густым и пышным. Такое качество особенно удобно для начинающих дачников. Если в первый год вы неудачно сформировали крону, уже к осени все ошибки закроются молодыми ветками, а весной вы придадите пузыреплоднику нужный контур.
Декоративные кусты используются для разных элементов дачного дизайна:
- устройства зеленых заборов;
- декорирования фонтанов, беседок, скульптур;
- создания композиций из нескольких растений или одиночных кустов;
- в качестве декоративного элемента на клумбах и цветниках.
Если спирея используется для живой изгороди, ждать окончания цветения нельзя: забор потеряет правильные очертания, во все стороны будут торчать молодые веточки, как иглы у ежа. Стрижку нужно провести ранней весной, пока не распустились почки. Примерно через месяц вы опять увидите торчащие веточки. Срезайте их своевременно, чтобы ограждение не стало бесформенным. Забор можно делать ровным, чередуя кусты с разной расцветкой листьев, а можно выполнить уступы, волны, башенки. Если дополнить пузыреплодник другими декоративными кустарниками или деревцами, простор для фантазии станет еще шире.
Из раскидистых видов, ветки которых сначала растут вверх, а концы опускаются вниз, можно создать густой широкий куст.
 Обрезайте нижние ветки примерно до 40 см от поверхности земли, у остальных побегов подравнивайте кончики. В зависимости от густоты побегов вы можете создать сплошной «зонтик» или оставить 4-5 стволов и получить фигурку, напоминающую осьминога. Для кроны такой формы хорошо подходит спирея калинолистная. На пустой лужайке хаотично расположите несколько кустиков, и участок приобретет оригинальный вид. Еще интереснее, если среди пониклых видов посадить растения с побегами, торчащими вверх.
Обрезайте нижние ветки примерно до 40 см от поверхности земли, у остальных побегов подравнивайте кончики. В зависимости от густоты побегов вы можете создать сплошной «зонтик» или оставить 4-5 стволов и получить фигурку, напоминающую осьминога. Для кроны такой формы хорошо подходит спирея калинолистная. На пустой лужайке хаотично расположите несколько кустиков, и участок приобретет оригинальный вид. Еще интереснее, если среди пониклых видов посадить растения с побегами, торчащими вверх.Совет
Посадите вокруг фонтана несколько кустов пузыреплодника калинолистного. Его раскидистые, поникшие на концах ветки будут повторять форму струй.
Когда куст станет достаточно густым, его кроне можно придать любую форму. Высокие округлые кусты можно сделать центром цветника или посадить около крыльца. Если собираетесь придать пузыреплоднику форму геометрической фигуры с прямыми гранями, будьте готовы к тому, что корректировать крону придется очень часто. Такая же проблема возникнет, если вы захотите сделать зеленые скульптуры: молодые побеги будут выбиваться за пределы контура то с одной, то с другой стороны, и секатор станет вашим постоянным инструментом.
Ландшафтные дизайнеры предлагают и другие варианты использования пузыреплодника, для каждого из них требуется своя форма кроны.
- Бордюр. Если вместо высокого зеленого забора устроить низкое ограждение, оно не закроет вид на участок, зато подчеркнет контур дорожек, выделит границу других сооружений.
- Альпийская горка. Для оформления этого объекта подходят карликовые сорта. Свисающие побеги создадут эффект кружевного покрывала.
- Массив. Если хотите, чтобы пузыреплодник требовал минимального ухода, посадите группу кустов вплотную друг к другу. Ровно подстриженный верх создаст эффект объемного зеленого ковра, уложенного на газон.
Иногда начинающие садоводы не выращивают на участке декоративные кустарники, потому что не умеют стричь их. На самом деле научиться этому можно за один сезон. Начните с пузыреплодника, его побеги растут очень быстро. Как у сказочного чудовища, на месте одной неправильно срезанной ветки вырастут 3 новых побега. На следующий год вы уже и сами не сможете заметить свою оплошность.
как правильно формировать, схема, видео
Содержимое
- 1 Можно ли обрезать пузыреплодник
- 2 Когда лучше обрезать пузыреплодник — весной или осенью
- 3 Виды обрезки пузыреплодника калинолистного
- 3.1 Омолаживающая обрезка пузыреплодника
- 3.2 Формирующая
- 3.3 Санитарная
- 4 Как формировать пузыреплодник
- 4.1 Каркасы
- 4.2 Живая изгородь
- 5 Как правильно обрезать пузыреплодник весной
- 6 Уход после весенней обрезки пузыреплодника
- 7 Заключение
Пузыреплодник калинолистный стал очень популярен в ландшафтном дизайне. К условиям выращивания кустарник неприхотлив. В течение всего сезона пузыреплодник сохраняет декоративность. Резные листья разного оттенка, пышные светлые соцветия и плоды интересной формы придают посадкам особый шарм. Обрезка пузыреплодника проводится для сохранения здоровья растения и придания кустам формы.
Можно ли обрезать пузыреплодник
Для создания живых изгородей или эффектных групповых композиций пузыреплодник подходит идеально. Крона до двух метров в диаметре с резными листьями быстро наращивается. Чтобы пузыреплодник калинолистный смотрелся ухоженным и сохранял выбранную форму, требуются формирующие стрижки по мере отрастания свежих побегов.
Неприхотливое растение хорошо зимует даже в холодном климате. Но невызревшие ветки могут пострадать от мороза или сломаться под гнетом снежного покрова. Такие побеги должны быть удалены.
Пузыреплодник к болезням устойчив. Но при нарушении правил ухода кустарник могут поражать некоторые вредители. Чтобы избежать гибели растения, требуется провести профилактические обрезки и санитарную обработку от болезней и вредителей.
Когда лучше обрезать пузыреплодник — весной или осенью
Обрезку пузыреплодника необходимо проводить несколько раз в год. Весной до начала цветения необходимо удалить все поврежденные или больные ветки. Это поможет кустарнику правильно развиваться, без трат времени и сил на восстановление.
Это поможет кустарнику правильно развиваться, без трат времени и сил на восстановление.
Когда пузыреплодник отцветет, обрезку проводят для формирования кроны. Иногда для этого требуется несколько мероприятий в течение сезона.
Осенью обрезку проводят для подготовки к зимней спячке.
Виды обрезки пузыреплодника калинолистного
Садоводы подразделяют виды обрезки пузыреплодника по целям, с которыми они проводятся.
Омолаживающая обрезка пузыреплодника
С годами кустарник стареет. Одревесневшие толстые ветки мешают дальнейшему развитию растения и портят его внешний вид. Первую обрезку для омоложения куста необходимо проводить по достижении пятилетнего возраста. Удалить требуется все толстые ветки полностью.
Формирующая
Обрезки для придания кустарнику красивой формы делают после цветения. Существует несколько форм стрижки. Но в большинстве случаев все зависит от фантазии садовода и умения правильно ухаживать за декоративным растением.
Санитарная
Стрижку пузыреплодника калинолистного с целью сохранения здоровья растения проводят весной и осенью. При санитарной обрезке необходимо удалить больные ветки и защитить кустарник от воздействия грибковой инфекции и вредителей.
При санитарной обрезке необходимо удалить больные ветки и защитить кустарник от воздействия грибковой инфекции и вредителей.
Как формировать пузыреплодник
Формирующую обрезку пузыреплодника весной лучше проводить после того, как кустарник отцветет.
Учитывая естественный рост и расположение веток пузыреплодника, его крону рекомендуют формировать в виде шара или фонтана.
Новички могут выбрать в питомнике уже сформированный шаром кустик для посадки в саду. В этом случае потребуется только поддерживать форму в течение сезона.
Самостоятельно формировку пузыреплодника следует начинать на второй год после посадки растения. Задачей первых стрижек является наращивание объема кроны на высоте от полуметра. Для этого следует укорачивать ветви наполовину, чтобы быстрее развивались молодые побеги. В дальнейшем обрезку ветвей делают на треть от длины наращивания.
Проще всего вырастить пузыреплодник в форме фонтана. С этой целью укорачивают нижние ветви по периметру, оставляя те, что растут из середины. «На глазок» обрезают излишки поросли, выступающей за видимые пределы запланированной фигуры. Получить «фонтан» из пузыреплодника можно уже на второй-третий год после посадки куста.
«На глазок» обрезают излишки поросли, выступающей за видимые пределы запланированной фигуры. Получить «фонтан» из пузыреплодника можно уже на второй-третий год после посадки куста.
Если у кустарника регулярно подравнивать вершину, то на фоне газона посадки из пузыреплодника будут смотреться пышным ковриком.
Невероятно много усилий потребуется, если садовод решит создать из пузыреплодника геометрически ровную фигуру. Для поддержания формы придется очень часто пользоваться секатором, чтобы быстро отрастающие ветки не портили вид фигуры. Различные формы обрезки выполняют по схемам.
Каркасы
Чтобы получить строгую форму куста в виде шара, можно использовать специальные каркасы. Приспособление устанавливают вблизи ствола и обрезают все ветви, которые выступают за отведенные пределы.
С помощью каркасов можно придать кустарнику любую форму. Такими устройствами пользуются дизайнеры для создания живых растительных скульптур в саду и получения топиария интересной формы.
При создания крупных парковых ансамблей из пузыреплодника, несколько кустов высаживают рядом. Стрижку проводят с учетом необходимых контуров фигуры.
Важно! Обрезку необходимо проводить качественными остро заточенными инструментами.
Живая изгородь
Формирующую обрезку живой изгороди начинают ранней весной. Не стоит дожидаться, пока куст отцветет. К этому времени изгородь может превратиться в растрепанную некрасивую посадку.
Формирование и поддержание аккуратного вида живой изгороди или бордюра нужно проводить регулярно, по мере отрастания поросли. Стрижку делают до 4 — 5 раз за сезон, чтобы сохранить форму зеленых насаждений.
Совет! Новичкам-садоводам рекомендуют использовать деревянные или проволочные каркасы на начальной стадии формирования бордюров.
Как правильно обрезать пузыреплодник весной
Ранней весной, когда почки еще только готовятся распуститься, нужно провести санитарную обрезку. Возможно, что не все побеги одинаково хорошо перезимовали. Тогда требуется удалить все поврежденные части ветвей у молодых кустов.
Тогда требуется удалить все поврежденные части ветвей у молодых кустов.
Ветви обрезают частично или полностью, в зависимости от состояния. Можно совместить санитарную обрезку с формирующей. Но ни в коем случае не следует оставлять поврежденные побеги ради сохранения формы куста.
Пузыреплодник легко переносит стрижку. Даже удаление большого количества ветвей восстановится достаточно быстро благодаря качественной подкормке и правильному уходу.
Более зрелые растения (после 5 лет) придется подвергнуть омолаживающей стрижке, чтобы старые ветви не препятствовали развитию молодой поросли.
Определить необходимость омолаживающей стрижки поможет наблюдение за состоянием куста:
- Если снизилось количество соцветий в период цветения.
- Цветы измельчали.
- Уменьшилась облиственность куста.
Старые ветви при омолаживающей обрезке пузыреплодника весной удаляют до самого основания, оставляя небольшой (около 5 — 7 см) пенек. Для работы можно использовать сучкорезы или специальные пилы. Действия должны быть точно рассчитаны, чтобы не повредить молодую поросль.
Действия должны быть точно рассчитаны, чтобы не повредить молодую поросль.
Чтобы кустарник не пострадал от инфекции, после обрезки толстых старых ветвей необходимо обработать места срезов противогрибковым препаратом и замазать садовым варом.
Важно! Формирование кустарника лучше проводить в вечернее время в сухую погоду.
Омолаживающие обрезки можно проводить не только по весне, но и осенью. Мероприятие проводят после опадания листвы. Важно учесть прогноз погоды, чтобы удаление старой поросли не привело к гибели куста от ранних морозов.
Старые ветви необходимо обязательно убрать и сжечь. Именно старая древесина часто становится рассадником инфекции. Многие вредители зимуют в растительных остатках, а затем, просыпаясь по весне, уничтожают посадки декоративных и плодовых культур.
Пример обрезки пузыреплодника весной детально показан на видео:
Уход после весенней обрезки пузыреплодника
Кустарник хорошо переносит обрезку и становится пышнее после удаления старых и больных частей. Но для наращивания молодой поросли ему требуется подкормка.
Но для наращивания молодой поросли ему требуется подкормка.
Чтобы растение получило все необходимые компоненты для развития, его необходимо подкормить органикой и минеральным комплексом. Вносят удобрения в виде раствора под корень.
Для подкормки взрослого куста стриженого пузыреплодника потребуется до 10 л питательной жидкости. Можно использовать коровяк в разведенном виде. На 10 литров воды 4 — 5 кг коровьего навоза необходимо настоять в течение нескольких часов. Литр получившейся жидкости нужно развести водой и полить куст. Специалисты советуют предварительно вылить под куст ведро воды, чтобы питательный раствор равномерно распределился и не повредил корневую систему растения.
Для восполнения минеральных компонентов после весенней обрезки можно использовать комплексные препараты. Нитроаммофоску разводят из расчета 30 г на 10 л. Калийную селитру добавляют вместе с мочевиной и коровяком. Для приготовления раствора потребуется по столовой ложке удобрений и около стакана раствора коровяка на ведро воды. Этой смеси будет достаточно для удобрения взрослого куста.
Этой смеси будет достаточно для удобрения взрослого куста.
Раны, полученные кустарником во время обрезки, могут стать воротами для инфекции. С целью защиты стоит обработать кусты стандартными комплексными средствами от вредителей и грибка.
Заключение
Обрезка пузыреплодника необходима даже в случае, если не планируется придавать кусту особую форму. Быстрое наращивание новой поросли постепенно угнетает рост кустарника. А его листва и цветы теряют декоративность, если не удалять старые и поврежденные ветви.
Механизм везикулярного транспорта — Клетка
Как видно из предыдущих разделов этой главы, транспортные везикулы играют центральную роль в перемещении молекул между различными окруженными мембраной компартментами секреторного пути. Как обсуждалось в главе 12, везикулы сходным образом участвуют в транспорте материалов, поглощаемых клеточной поверхностью. Таким образом, везикулярный транспорт является основной клеточной активностью, ответственной за молекулярный транспорт между множеством специфических компартментов, окруженных мембраной. Таким образом, избирательность такого транспорта является ключом к поддержанию функциональной организации клетки. Например, лизосомальные ферменты должны транспортироваться специфически из аппарата Гольджи в лизосомы, а не на плазматическую мембрану или в ЭПР. Некоторые из сигналов, которые нацеливают белки на специфические органеллы, такие как лизосомы, обсуждались ранее в этой главе. Эти белки транспортируются внутри везикул, поэтому специфичность транспорта основана на селективной упаковке предназначенного груза в везикулы, которые распознают и сливаются только с соответствующей мембраной-мишенью. Из-за центральной важности везикулярного транспорта для организации эукариотических клеток понимание молекулярных механизмов, которые контролируют упаковку, почкование и слияние пузырьков, является основной областью исследований в области клеточной биологии.
Таким образом, избирательность такого транспорта является ключом к поддержанию функциональной организации клетки. Например, лизосомальные ферменты должны транспортироваться специфически из аппарата Гольджи в лизосомы, а не на плазматическую мембрану или в ЭПР. Некоторые из сигналов, которые нацеливают белки на специфические органеллы, такие как лизосомы, обсуждались ранее в этой главе. Эти белки транспортируются внутри везикул, поэтому специфичность транспорта основана на селективной упаковке предназначенного груза в везикулы, которые распознают и сливаются только с соответствующей мембраной-мишенью. Из-за центральной важности везикулярного транспорта для организации эукариотических клеток понимание молекулярных механизмов, которые контролируют упаковку, почкование и слияние пузырьков, является основной областью исследований в области клеточной биологии.
Экспериментальные подходы к пониманию везикулярного транспорта
Прогресс в выяснении молекулярных механизмов везикулярного транспорта был достигнут благодаря трем различным экспериментальным подходам: (1) выделение дрожжевых мутантов, дефектных в транспорте и сортировке белков; (2) восстановление везикулярного транспорта в бесклеточных системах; и (3) биохимический анализ синаптических пузырьков, отвечающих за регулируемую секрецию нейротрансмиттеров нейронами. Каждая из этих экспериментальных систем имеет определенные преимущества для понимания конкретных аспектов транспортного процесса. Наиболее важным, однако, является тот факт, что результаты всех трех направлений исследований сошлись, указывая на то, что сходные молекулярные механизмы регулируют секрецию в таких разных клетках, как дрожжи и нейроны млекопитающих.
Каждая из этих экспериментальных систем имеет определенные преимущества для понимания конкретных аспектов транспортного процесса. Наиболее важным, однако, является тот факт, что результаты всех трех направлений исследований сошлись, указывая на то, что сходные молекулярные механизмы регулируют секрецию в таких разных клетках, как дрожжи и нейроны млекопитающих.
Как и в других областях клеточной биологии, дрожжи оказались полезными при изучении секреторного пути, поскольку они легко поддаются генетическому анализу. В частности, Randy Schekman и его коллеги первыми выделили дрожжевые мутанты, дефектные по везикулярному транспорту. К ним относятся мутанты, дефектные на различных стадиях секреции белка (мутанты sec ), мутанты, неспособные транспортировать белки в вакуоли, и мутанты, неспособные сохранять резидентные белки ER. Выделение таких мутантов у дрожжей привело непосредственно к молекулярному клонированию и анализу соответствующих генов, тем самым идентифицировав ряд белков, участвующих в различных стадиях секреторного пути. Например, ранее в этой главе обсуждалась роль Sec61 как основного компонента канала транслокации белка в эндоплазматическом ретикулуме.
Например, ранее в этой главе обсуждалась роль Sec61 как основного компонента канала транслокации белка в эндоплазматическом ретикулуме.
Биохимические исследования везикулярного транспорта с использованием реконструированных систем дополнили эти генетические исследования и позволили напрямую выделить транспортные белки из клеток млекопитающих. Первая бесклеточная транспортная система была разработана Джеймсом Ротманом и его коллегами, которые проанализировали транспорт белков между отделами аппарата Гольджи (14). В экспериментальном плане использовалась мутантная клеточная линия млекопитающих, в которой отсутствовал фермент, необходимый для переноса остатков N -ацетилглюкозамина в N -связанный олигосахарид на ранней стадии его модификации в аппарате Гольджи (см. ). Следовательно, в гликопротеинах, продуцируемых этой мутантной клеточной линией, отсутствовали дополнительные единицы N -ацетилглюкозамина. Однако если стопки Гольджи, выделенные из мутантной клеточной линии, инкубировали со стопками, выделенными из нормальных клеток, к гликопротеинам, синтезируемым мутантными клетками, добавлялись остатки N -ацетилглюкозамина. Множество экспериментов установило, что это происходит в результате везикулярного транспорта белков из стопок Гольджи мутантной клеточной линии в стопки Гольджи нормальных клеток, поэтому добавление N -ацетилглюкозамин представляет собой легко определяемый маркер везикулярного транспорта в этой реконструированной системе. Подобные восстановленные системы были разработаны для анализа транспорта между другими компартментами, включая транспорт из ER в аппарат Гольджи и транспорт из аппарата Гольджи в секреторные везикулы, вакуоли и плазматическую мембрану. Разработка этих систем in vitro позволила провести биохимические исследования процесса транспорта и функциональный анализ белков, идентифицированных в результате мутаций у дрожжей, а также прямое выделение некоторых белков, участвующих в почковании и слиянии пузырьков.
Множество экспериментов установило, что это происходит в результате везикулярного транспорта белков из стопок Гольджи мутантной клеточной линии в стопки Гольджи нормальных клеток, поэтому добавление N -ацетилглюкозамин представляет собой легко определяемый маркер везикулярного транспорта в этой реконструированной системе. Подобные восстановленные системы были разработаны для анализа транспорта между другими компартментами, включая транспорт из ER в аппарат Гольджи и транспорт из аппарата Гольджи в секреторные везикулы, вакуоли и плазматическую мембрану. Разработка этих систем in vitro позволила провести биохимические исследования процесса транспорта и функциональный анализ белков, идентифицированных в результате мутаций у дрожжей, а также прямое выделение некоторых белков, участвующих в почковании и слиянии пузырьков.
Рисунок 9.30
Восстановленный везикулярный транспорт. Наборы Гольджи, приготовленные из инфицированной вирусом мутантной клеточной линии, неспособной катализировать добавление N -ацетилглюкозамина к N -связанным олигосахаридам, смешивают с наборами Гольджи из нормальной клеточной линии. Потому что мутант (подробнее…)
Потому что мутант (подробнее…)
Критическое понимание молекулярных механизмов везикулярного транспорта также было получено в результате изучения синаптической передачи в нейронах, которая представляет собой высокоспециализированную форму регулируемой секреции. Синапс — это соединение нейрона с другой клеткой, которая может быть либо другим нейроном, либо эффектором, например мышечной клеткой. Информация передается через синапс с помощью химических нейротрансмиттеров, таких как ацетилхолин, которые хранятся в синаптических пузырьках. Стимуляция передающего нейрона запускает слияние синаптических пузырьков с плазматической мембраной, вызывая высвобождение нейротрансмиттеров и стимулируя постсинаптический нейрон или эффекторную клетку. Синаптических везикул чрезвычайно много в головном мозге, что позволяет очищать их в больших количествах для биохимического анализа. Некоторые из белков, выделенных из синаптических везикул, тесно связаны с белками, которые, как было показано, играют критическую роль в везикулярном транспорте в экспериментах по генетике и восстановлению дрожжей, поэтому биохимический анализ этих белков дал важную информацию о молекулярных механизмах слияния везикул.
Белки оболочки и отпочкование везикул
Первым этапом везикулярного транспорта является образование везикул путем отпочкования от мембраны. Цитоплазматические поверхности транспортных везикул покрыты белками, и, по-видимому, сборка этих белковых оболочек управляет почкованием везикул путем искажения конформации мембраны. Были охарактеризованы три вида покрытых оболочкой везикул, которые, по-видимому, функционируют в различных типах везикулярного транспорта. Первыми были описаны покрытые клатрином везикулы , которые отвечают за поглощение внеклеточных молекул из плазматической мембраны путем эндоцитоза (см. главу 12), а также за транспорт молекул из транс сети Гольджи в лизосомы. Два других типа покрытых везикул были идентифицированы как отпочковавшиеся от ER и комплекса Гольджи. Эти везикулы называются непокрытыми клатрином или покрытыми COP везикулами (COP обозначает белок оболочки). Один класс этих везикул (везикулы, покрытые COPII) отпочковываются от ER и переносят свой груз вперед по секреторному пути к аппарату Гольджи. Напротив, COPI-покрытые везикулы отпочковываются от промежуточного компартмента ER-Golgi или аппарата Гольджи и функционируют в путях извлечения, которые служат для сохранения резидентных белков в Golgi и ER. Например, везикулы, покрытые COPI, транспортируют резидентные белки ER, отмеченные сигналами поиска KDEL или KKXX, обратно в ER из промежуточного компартмента ER-Golgi или цис Сеть Гольджи.
Напротив, COPI-покрытые везикулы отпочковываются от промежуточного компартмента ER-Golgi или аппарата Гольджи и функционируют в путях извлечения, которые служат для сохранения резидентных белков в Golgi и ER. Например, везикулы, покрытые COPI, транспортируют резидентные белки ER, отмеченные сигналами поиска KDEL или KKXX, обратно в ER из промежуточного компартмента ER-Golgi или цис Сеть Гольджи.
Оболочки везикул, покрытых клатрином, состоят из белковых комплексов двух типов, клатрина и адапторных белков, которые собираются на цитозольной стороне мембран (). Clathrin играет структурную роль, собираясь в решетчатую структуру, подобную корзине, которая искажает мембрану и управляет почкованием везикул. Связывание клатрина с мембранами опосредовано вторым классом белков, называемых адапторными белками. Различные адапторные белки ответственны за сборку покрытых клатрином везикул на плазматической мембране и в транс сети Гольджи, и именно адапторные белки участвуют в выборе специфических молекул для включения в везикулы. Напр., адапторный белок АР-1, участвующий в отпочковании от сети Гольджи trans , связывается с цитозольной частью маннозо-6-фосфатного рецептора, тем самым направляя белки, предназначенные для лизосом, в покрытые клатрином везикулы.
Напр., адапторный белок АР-1, участвующий в отпочковании от сети Гольджи trans , связывается с цитозольной частью маннозо-6-фосфатного рецептора, тем самым направляя белки, предназначенные для лизосом, в покрытые клатрином везикулы.
Рисунок 9.31
Включение лизосомальных белков в везикулы, покрытые клатрином. Белки, нацеленные на лизосомы, помечены маннозо-6-фосфатами, которые связываются с маннозо-6-фосфатными рецепторами в транс Сеть Гольджи. Рецепторы маннозо-6-фосфата охватывают Гольджи (подробнее…)
Оболочки COPI- и COPII-покрытых везикул состоят из различных белковых комплексов, которые функционируют аналогично клатрину и адапторным белкам при отпочковывании везикул. Интересно, что компоненты оболочки COPI взаимодействуют с мотивом KKXX, ответственным за извлечение белков ER из аппарата Гольджи, что согласуется с ролью COPI-покрытых везикул в рециркуляции от Гольджи к ER.
Сборка оболочек везикул также требует GTP-связывающих белков, которые, по-видимому, регулируют связывание белков оболочки с мембраной. Для отпочкования везикул, покрытых клатрином, и везикул, покрытых COPI, из комплекса Гольджи требуется GTP-связывающий белок, называемый ARF (фактор рибозилирования АДФ), в то время как для отпочковывания везикул, покрытых COPII, от ER требуется отдельный GTP-связывающий белок, называемый Сар1. Роль этих белков иллюстрируется функцией ARF в сборке везикул, покрытых COPI (10). Первым этапом образования везикул является ассоциация ARF, связанного с GDP, с мембраной Гольджи. Затем белки в мембране Гольджи стимулируют обмен GDP, связанного с ARF, на GTP, а белки оболочки COPI связываются с комплексом ARF/GTP. Затем за сборкой оболочки следует деформация мембраны и почкование пузырьков. Затем ARF гидролизует свой связанный GTP, что приводит к превращению ARF в состояние, связанное с GDP, и диссоциации белков оболочки от мембраны везикул.
Для отпочкования везикул, покрытых клатрином, и везикул, покрытых COPI, из комплекса Гольджи требуется GTP-связывающий белок, называемый ARF (фактор рибозилирования АДФ), в то время как для отпочковывания везикул, покрытых COPII, от ER требуется отдельный GTP-связывающий белок, называемый Сар1. Роль этих белков иллюстрируется функцией ARF в сборке везикул, покрытых COPI (10). Первым этапом образования везикул является ассоциация ARF, связанного с GDP, с мембраной Гольджи. Затем белки в мембране Гольджи стимулируют обмен GDP, связанного с ARF, на GTP, а белки оболочки COPI связываются с комплексом ARF/GTP. Затем за сборкой оболочки следует деформация мембраны и почкование пузырьков. Затем ARF гидролизует свой связанный GTP, что приводит к превращению ARF в состояние, связанное с GDP, и диссоциации белков оболочки от мембраны везикул.
Рис. 9.32
Роль ARF в формировании везикул, покрытых COP. ARF чередуется между состояниями, связанными с GTP, и состояниями, связанными с GDP.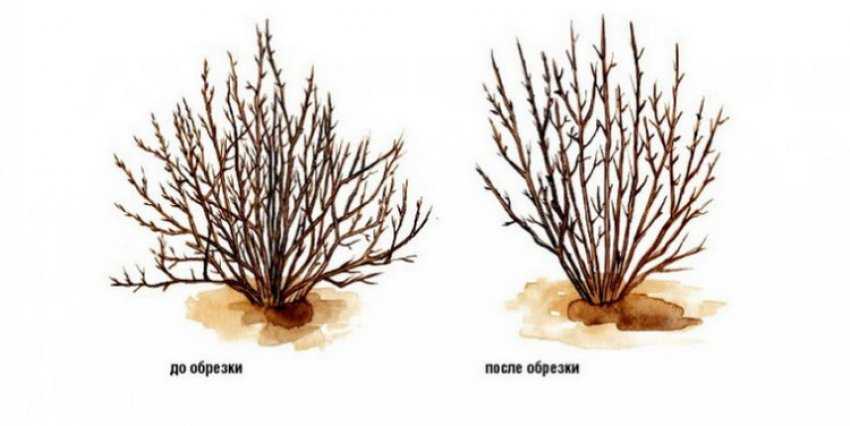 При связывании с GDP ARF связывается с мембраной транс сети Гольджи, где факторы обмена гуаниновых нуклеотидов (GEF) способствуют обмену (подробнее…) включает в себя два типа событий. Во-первых, транспортная везикула должна специфически распознавать правильную мембрану-мишень; например, везикула, несущая лизосомальные ферменты, должна доставлять свой груз только к лизосомам. Во-вторых, везикула и мембраны-мишени должны сливаться, тем самым доставляя содержимое везикулы к органелле-мишени. Исследования, проведенные за последние несколько лет, привели к разработке модели слияния везикул, в которой специфическое распознавание между везикулой и ее мишенью опосредуется взаимодействием между уникальными парами трансмембранных белков с последующим слиянием фосфолипидных бислоев везикулы и мембраны-мишени. .
При связывании с GDP ARF связывается с мембраной транс сети Гольджи, где факторы обмена гуаниновых нуклеотидов (GEF) способствуют обмену (подробнее…) включает в себя два типа событий. Во-первых, транспортная везикула должна специфически распознавать правильную мембрану-мишень; например, везикула, несущая лизосомальные ферменты, должна доставлять свой груз только к лизосомам. Во-вторых, везикула и мембраны-мишени должны сливаться, тем самым доставляя содержимое везикулы к органелле-мишени. Исследования, проведенные за последние несколько лет, привели к разработке модели слияния везикул, в которой специфическое распознавание между везикулой и ее мишенью опосредуется взаимодействием между уникальными парами трансмембранных белков с последующим слиянием фосфолипидных бислоев везикулы и мембраны-мишени. .
Белки, участвующие в слиянии везикул, были первоначально идентифицированы в лаборатории Джеймса Ротмана путем биохимического анализа восстановленных систем везикулярного транспорта из клеток млекопитающих (см. ). Анализ белков, участвующих в слиянии везикул в этих системах, привел Ротмана и его коллег к предложению общей модели, названной гипотезой SNARE , в которой слияние везикул опосредовано взаимодействиями между специфическими парами белков, называемых SNARE, на везикуле и мембраны-мишени (v-SNARE и t-SNARE соответственно) (). Эта гипотеза была подтверждена идентификацией SNARE, которые присутствовали на синаптических везикулах, и обнаружением мутантов дрожжевой секреции, которые, по-видимому, кодируют SNARE, необходимые для различных событий транспорта везикул. Напр., для транспорта из ER в аппарат Гольджи у дрожжей требуются специфические SNARE, расположенные как на мембранах везикул, так и на мембранах-мишенях. Образование комплексов между v-SNARE на везикулах и t-SNARE на мембранах-мишенях затем приводит к слиянию мембран с помощью механизмов, которые еще предстоит полностью понять.
). Анализ белков, участвующих в слиянии везикул в этих системах, привел Ротмана и его коллег к предложению общей модели, названной гипотезой SNARE , в которой слияние везикул опосредовано взаимодействиями между специфическими парами белков, называемых SNARE, на везикуле и мембраны-мишени (v-SNARE и t-SNARE соответственно) (). Эта гипотеза была подтверждена идентификацией SNARE, которые присутствовали на синаптических везикулах, и обнаружением мутантов дрожжевой секреции, которые, по-видимому, кодируют SNARE, необходимые для различных событий транспорта везикул. Напр., для транспорта из ER в аппарат Гольджи у дрожжей требуются специфические SNARE, расположенные как на мембранах везикул, так и на мембранах-мишенях. Образование комплексов между v-SNARE на везикулах и t-SNARE на мембранах-мишенях затем приводит к слиянию мембран с помощью механизмов, которые еще предстоит полностью понять.
Рисунок 9.33
Слияние везикул. Слияние везикул опосредуется связыванием между специфическими парами v-SNARE и t-SNARE на мембранах везикул и мишенях соответственно. Rab GTP-связывающие белки необходимы для облегчения образования комплексов v-SNARE/t-SNARE. Далее (подробнее…)
Rab GTP-связывающие белки необходимы для облегчения образования комплексов v-SNARE/t-SNARE. Далее (подробнее…)
В дополнение к SNARE для слияния везикул требуются как минимум два других типа белков. Белки Rab представляют собой семейство малых GTP-связывающих белков, которые родственны белкам Ras, которые обсуждались в главе 7. Было идентифицировано более 30 различных белков Rab, и показано, что они функционируют в специфических процессах транспорта везикул. Они могут функционировать на нескольких этапах доставки везикул, включая взаимодействие с SNARE для регуляции и облегчения образования комплексов v-SNARE/t-SNARE.
После образования комплексов между комплементарными SNARE и слияния мембран для завершения процесса транспорта везикул необходим комплекс из двух дополнительных белков (комплекс NSF/SNAP). Белки NSF/SNAP рекрутируются на мембраны после образования комплексов v-SNARE/t-SNARE, и они не требуются напрямую ни для спаривания везикул/мишеней, ни для слияния спаренных мембран. Вместо этого белки NSF/SNAP действуют после слияния мембран, чтобы разобрать комплекс SNARE, тем самым позволяя повторно использовать SNARE для последующих раундов транспорта везикул.
Вместо этого белки NSF/SNAP действуют после слияния мембран, чтобы разобрать комплекс SNARE, тем самым позволяя повторно использовать SNARE для последующих раундов транспорта везикул.
11.6: Везикулярный транспорт — Биология LibreTexts
- Последнее обновление
- Сохранить как PDF
- Идентификатор страницы
- 16161
- Э. В. Вонг
- Axolotl Academica Publishing
В дополнение к процессингу белков, ER и Golgi также заботятся о некоторых типах транспорта белков. Везикулы (по существу, связанные с мембраной пузырьки) отщепляются от ЭПР, аппарата Гольджи и других мембранных органелл, унося с собой любые растворимые молекулы, находящиеся внутри заключенной жидкости, а также любые молекулы, встроенные в этот участок мембраны. Затем эти везикулы подхватывают молекулярный двигатель, такой как кинезин или миозин, и путешествуют по цитоскелету, пока не стыкуются в соответствующем месте и не сливаются с мембраной-мишенью или органеллой. В общем, везикулы перемещаются из ER в цис-Гольджи, из цис-в медиальный аппарат Гольджи, из медиального в транс-Гольджи и из транс-Гольджи в плазматическую мембрану или другие компартменты. Хотя большая часть движения происходит в этом направлении, есть также везикулы, которые перемещаются обратно от аппарата Гольджи к ЭПР, неся белки, которые должны были оставаться в ЭПР (например, PDI) и были случайно скоплены внутри везикулы.
Затем эти везикулы подхватывают молекулярный двигатель, такой как кинезин или миозин, и путешествуют по цитоскелету, пока не стыкуются в соответствующем месте и не сливаются с мембраной-мишенью или органеллой. В общем, везикулы перемещаются из ER в цис-Гольджи, из цис-в медиальный аппарат Гольджи, из медиального в транс-Гольджи и из транс-Гольджи в плазматическую мембрану или другие компартменты. Хотя большая часть движения происходит в этом направлении, есть также везикулы, которые перемещаются обратно от аппарата Гольджи к ЭПР, неся белки, которые должны были оставаться в ЭПР (например, PDI) и были случайно скоплены внутри везикулы.
Формирование везикул зависит от белков оболочки, которые при соответствующих условиях самособираются в сферические клетки. Когда они связаны с трансмембранными белками, они также могут тянуть прикрепленную мембрану, придавая ей сферическую форму. Основными типами белков оболочки, используемых при образовании везикул, являются COPII, COPI и клатрин.
Когда они связаны с трансмембранными белками, они также могут тянуть прикрепленную мембрану, придавая ей сферическую форму. Основными типами белков оболочки, используемых при образовании везикул, являются COPII, COPI и клатрин.
COPII образуют везикулы, которые перемещаются от ER к аппарату Гольджи. Белки оболочки COPI используются между частями аппарата Гольджи, а также для формирования везикул, идущих от аппарата Гольджи обратно в ER. Наконец, клатрин используется для образования везикул, покидающих аппарат Гольджи для плазматической мембраны, а также для везикул, образующихся из плазматической мембраны для эндоцитоза.
Рисунок \(\PageIndex{17}\). Клатрин. (A) клатрин связывается с адапторными белками, которые связаны с трансмембранными грузовыми рецепторами, связывая мембрану с клатрином. (B) Один клатриновый трискелион состоит из трех тяжелых цепей и трех легких цепей. (c) Трискелионы самостоятельно собираются в примерно сферическую конструкцию без необходимости какой-либо дополнительной энергии или ферментов.
Клатрин (Рисунок \(\PageIndex{17}\)) лучше всего описан из трех, а везикулярные покровы состоят из композиций клатриновых трискелионов (от греческого, что означает трехногий). Каждый трискелион состоит из трех тяжелых цепей, соединенных вместе на С-конце, и трех легких цепей, по одной связанной с каждой тяжелой цепью. Тяжелые цепи разных трискелионов взаимодействуют по длине их «ног» тяжелой цепи, создавая очень прочную конструкцию. Легкие цепи не нужны для образования везикул, и считается, что они помогают предотвратить случайные взаимодействия молекул клатрина в цитоплазме.
Существует значительное сходство между механизмами образования везикул с использованием этих разных белков оболочки, начиная с рекрутирования ARF1 (ARF обозначает фактор рибозилирования ADP, который здесь не имеет ничего общего с его функцией) на мембрану. Для этого требуется ARNO-облегченный обмен GTP на GDP (ARNO — это открыватель сайта связывания нуклеотидов ARF). Как только ARF1 связывается с GTP, конформационные изменения обнаруживают N-концевую миристоильную группу, которая внедряется в мембрану. И COPI, и покрытые клатрином везикулы используют ARF1 и ARNO, но COPII использует сходные белки, называемые Sar1p и Sec12p.
И COPI, и покрытые клатрином везикулы используют ARF1 и ARNO, но COPII использует сходные белки, называемые Sar1p и Sec12p.
Рисунок \(\PageIndex{18}\). Везикулы, покрытые КС
ARF1 (или Sar1p) используется для рекрутирования адапторных белков, которые связываются с «хвостовым» концом мембраносвязанных рецепторных белков. Деловой конец этих рецепторов связывается с молекулами груза, которые необходимо упаковать в везикулу. Адаптерные белки действуют как связующее звено между мембраной (через рецепторы) и белками оболочки. Для клатрина адапторными белками являются AP1 для везикул, происходящих от транс-Гольджи, и AP2 для эндоцитарных везикул. Для везикул COPI приблизительными гомологами являются β-, γ-, δ- и ζ-COP, тогда как в системе COPII используются Sec23p и Sec24p.
Наконец, адаптеры связаны с собственно белками оболочки: клатрином, α- или ε-COP, Sec13p и Sec31p. Общим для всех этих белков является то, что они спонтанно (т. е. без каких-либо затрат энергии) самособираются в клетки, подобные сферическим структурам. Под электронным микроскопом везикулы, покрытые клатрином, более четко очерчены, а гексагональные и пятиугольные формы, ограниченные клатриновыми субъединицами, придают везикулам вид «футбольного мяча». Везикулы, покрытые коаламером COP, выглядят гораздо более нечеткими под действием ЭМ.
Под электронным микроскопом везикулы, покрытые клатрином, более четко очерчены, а гексагональные и пятиугольные формы, ограниченные клатриновыми субъединицами, придают везикулам вид «футбольного мяча». Везикулы, покрытые коаламером COP, выглядят гораздо более нечеткими под действием ЭМ.
Все три типа белков оболочки везикул обладают способностью спонтанно связываться в сферическую конструкцию, но только везикула, покрытая COPI и COPII, также спонтанно «отщипывает» мембрану, чтобы освободить везикулу от ее исходной мембраны. Везикулы, покрытые клатрином, требуют внешнего механизма для высвобождения везикул (рис. \(\PageIndex{19}\)).
Рисунок \(\PageIndex{19}\). Мономеры динамина, каждый из которых представляет собой ГТФазу, полимеризуются вокруг шейки везикулы. Когда GTP гидролизуется, динаминовая «петля» затягивается и пережимает везикулу. Когда везикула почти завершена, остается небольшая ножка или шейка мембраны, которая соединяет везикулу с мембраной. Вокруг этого стебля динамические молекулы GTP собираются в кольцо/спираль. Молекулы динамина представляют собой глобулярные ГТФазы, которые сокращаются при гидролизе ГТФ. Когда они связываются вокруг ножки везикулы, каждый белок динамина сокращается с комбинированным эффектом сжатия ножки настолько, что мембрана сжимается вместе, изолируя и освобождая везикулу от исходной мембраны.
Молекулы динамина представляют собой глобулярные ГТФазы, которые сокращаются при гидролизе ГТФ. Когда они связываются вокруг ножки везикулы, каждый белок динамина сокращается с комбинированным эффектом сжатия ножки настолько, что мембрана сжимается вместе, изолируя и освобождая везикулу от исходной мембраны.
Хотя липиды и мембраны обсуждались в главе 4, мы забыли обсудить место их синтеза у эукариот. Как показано на рисунке \(\PageIndex{20}\), синтез определенных типов липидов является сегрегированным и исключительным. Глицерофосфолипиды в основном образуются в эндоплазматическом ретикулуме, хотя они также производятся в митохондриях и пероксисомах. Напротив, сфинголипиды не вырабатываются в ER (хотя их церамидные предшественники) у млекопитающих необходимые ферменты обнаруживаются в просвете цис- и медиального аппарата Гольджи. Имеются доказательства антероградного и ретроградного везикулярного движения между различными компартментами Гольджи и ER, что теоретически может указывать на перераспределение типов липидов. Однако сфинголипиды склонны к агрегации в липидные рафты и, по-видимому, более сконцентрированы в антероградно движущихся везикулах.
Имеются доказательства антероградного и ретроградного везикулярного движения между различными компартментами Гольджи и ER, что теоретически может указывать на перераспределение типов липидов. Однако сфинголипиды склонны к агрегации в липидные рафты и, по-видимому, более сконцентрированы в антероградно движущихся везикулах.
Белки оболочки отрываются вскоре после высвобождения везикул. Для клатрина в этом процессе участвует Hsc70, АТФаза. Однако для везикул, покрытых COPI или COPII, гидролиз GTP на ARF/Sar1p, по-видимому, ослабляет сродство белка оболочки к адаптерам и инициирует снятие покрытия. Активатор GTPase представляет собой ARF GAP (или Sec23p) и является неотъемлемой частью оболочки COP I (или II).
Везикулы несут две категории грузов: растворимые белки и трансмембранные белки. Из растворимых белков некоторые поглощаются везикулами благодаря связыванию с рецептором. Другие белки просто оказываются поблизости и захватываются по мере формирования везикулы. Иногда поглощается белок, которого не должно было быть; например, PDI может быть заключен в везикулу, формирующуюся из ER. У него мало функций в аппарате Гольджи, и он необходим в отделении неотложной помощи, так что же с ним происходит? К счастью, PDI и многие другие белки ER имеют С-концевую сигнальную последовательность, KDEL (лизин-аспарагиновая кислота-глутаминовая кислота-лейцин), которая кричит: «Я принадлежу ER». Эта последовательность распознается рецепторами KDEL внутри аппарата Гольджи, и связывание белков KDEL с рецепторами запускает образование везикул, чтобы отправить их обратно в ER.
У него мало функций в аппарате Гольджи, и он необходим в отделении неотложной помощи, так что же с ним происходит? К счастью, PDI и многие другие белки ER имеют С-концевую сигнальную последовательность, KDEL (лизин-аспарагиновая кислота-глутаминовая кислота-лейцин), которая кричит: «Я принадлежу ER». Эта последовательность распознается рецепторами KDEL внутри аппарата Гольджи, и связывание белков KDEL с рецепторами запускает образование везикул, чтобы отправить их обратно в ER.
У секреторных везикул особая проблема с растворимым грузом. Если бы везикула полагалась просто на включение в нее белков во время процесса формирования, было бы трудно получить высокие концентрации этих белков. Многие секретируемые белки необходимы организму быстро и в значительных количествах, поэтому в транс-Гольджи имеется механизм агрегации секреторных белков. Механизм использует агрегирующие белки, такие как секретогранин II и хромогранин B, которые объединяют целевые белки в большие концентрированные гранулы. Эти гранулины лучше всего работают в среде транс-Гольджи с низким pH и высоким содержанием Ca 9.0149 2+ , поэтому, когда везикула высвобождает свое содержимое за пределы клетки, более высокий pH и более низкий Ca 2+ разрушают агрегаты, чтобы высвободить отдельные белки.
Эти гранулины лучше всего работают в среде транс-Гольджи с низким pH и высоким содержанием Ca 9.0149 2+ , поэтому, когда везикула высвобождает свое содержимое за пределы клетки, более высокий pH и более низкий Ca 2+ разрушают агрегаты, чтобы высвободить отдельные белки.
Существует постоянное изменение pH во время созревания аппарата Гольджи, так что по мере перехода от ER к аппарату Гольджи каждый компартмент имеет все более низкий (более кислый) люменальный pH.
Наконец, возникает вопрос о нацеливании на везикулы. Везикулы гораздо менее полезны, если их бросить в молекулярный грузовой поезд и высадить наугад. Следовательно, существует механизм стыковки, который требует соответствия белка v-SNARE на цитоплазматической поверхности везикул и t-SNARE на цитоплазматической поверхности мембраны-мишени. Слияние пузырька с мембраной происходит только при наличии совпадения. В противном случае везикула не сможет слиться и прикрепится к другому молекулярному мотору, чтобы направиться к другому, надеюсь правильному, месту назначения. Этому процессу способствуют связывающие белки, которые первоначально вступают в контакт с входящей везикулой и приближают ее достаточно близко к мишени, чтобы проверить взаимодействие с белком SNARE. Затем другие белки на везикулах и мембранах-мишенях взаимодействуют, и, если SNARE совпадают, могут помочь «втянуть» везикулу в мембрану-мишень, после чего мембраны сливаются. Важным практическим правилом для понимания везикулярного слияния, а также направленности мембранных белков и липидов является то, что сторона мембраны, обращенная к цитоплазме, всегда будет обращена к цитоплазме. Следовательно, белок, который в конечном итоге обнаруживается на внешней поверхности клеточной мембраны, с самого начала будет вставлен в люменальную поверхность мембраны ER.
Этому процессу способствуют связывающие белки, которые первоначально вступают в контакт с входящей везикулой и приближают ее достаточно близко к мишени, чтобы проверить взаимодействие с белком SNARE. Затем другие белки на везикулах и мембранах-мишенях взаимодействуют, и, если SNARE совпадают, могут помочь «втянуть» везикулу в мембрану-мишень, после чего мембраны сливаются. Важным практическим правилом для понимания везикулярного слияния, а также направленности мембранных белков и липидов является то, что сторона мембраны, обращенная к цитоплазме, всегда будет обращена к цитоплазме. Следовательно, белок, который в конечном итоге обнаруживается на внешней поверхности клеточной мембраны, с самого начала будет вставлен в люменальную поверхность мембраны ER.

Более конкретно, когда везикула приближается к мембране-мишени, связывающий белок Rab-GTP, который связан с мембраной-мишенью через двойной геранилгеранил-липидный хвост, неплотно связывается с везикулой и удерживает ее вблизи мембраны-мишени, давая ЛОВУШКИ шанс работать. У v-SNARE и t-SNARE теперь есть возможность взаимодействовать и проверять соответствие. Недавно SNARE были переименованы в R-SNARE и Q-SNARE соответственно на основе консервативных остатков аргинина и глутамина. В дополнение к этим двум первичным SNARE участвует по крайней мере еще одна SNARE, вместе образующая пучок из четырех α-спиралей (четырех, а не трех, потому что, по крайней мере, в наиболее изученном примере одна из SNARE загибается так, что две из его альфа-спиральных доменов участвуют во взаимодействии.Четыре спирали закручиваются вокруг друг друга, и считается, что при этом они стягивают везикулу и мембрану-мишень вместе.
Столбнячный токсин, тетаноспазмин, который высвобождается бактериями Clostridium tetani , вызывает спазмы, воздействуя на нервные клетки и предотвращая высвобождение нейромедиаторов. Механизм этого заключается в том, что он расщепляет синаптобревин, белок SNARE, так что синаптические пузырьки не могут сливаться с клеточной мембраной. Ботулинический токсин из Clostridium botulinum также действует на SNARE, предотвращая слияние пузырьков и высвобождение нейротрансмиттеров, хотя он нацелен на разные нейроны и поэтому имеет противоположный эффект: столбняк вызывается предотвращением высвобождения ингибирующих нейротрансмиттеров, тогда как ботулизм вызывается предотвращением высвобождение возбуждающих нейротрансмиттеров.
Механизм этого заключается в том, что он расщепляет синаптобревин, белок SNARE, так что синаптические пузырьки не могут сливаться с клеточной мембраной. Ботулинический токсин из Clostridium botulinum также действует на SNARE, предотвращая слияние пузырьков и высвобождение нейротрансмиттеров, хотя он нацелен на разные нейроны и поэтому имеет противоположный эффект: столбняк вызывается предотвращением высвобождения ингибирующих нейротрансмиттеров, тогда как ботулизм вызывается предотвращением высвобождение возбуждающих нейротрансмиттеров.
Эта страница под названием 11.6: Vesicular Transport распространяется под лицензией CC BY-NC-SA 3.0 и была создана, изменена и/или курирована Э. В. Вонгом посредством исходного контента, который был отредактирован в соответствии со стилем и стандартами платформы LibreTexts; подробная история редактирования доступна по запросу.
- Наверх
- Была ли эта статья полезной?
- Тип изделия
- Раздел или страница
- Автор
- Э.
