Морфологическое описание акация белая: Робиния — PharmSpravka
Робиния — PharmSpravka
Flores Robiniae pseudacaciae — цветки робинии лжеакации
Робиния лжеакация — Robinia pseudacacia L.
Семейство бобовые — Fabaceae (Leguminosae)
Подсемейстов Faboideae
Другие названия:
— белая акация
Ботаническая характеристика. Робиния лжеакация — листопадное дерево до 30-35 м высоты. Листья очередные непарноперистосложные, 11-24 см длины с продолговатыми или эллиптическими заостренными листочками 1,5-4,5 см длины и 0,7-2 см ширины.
Разводится в садах, парках в европейской части России, на Кавказе, Дальнем Востоке России, в Центральной Азии. Родина — Северная Америка.
Химический состав. В цветках содержится эфирное масло, в состав которого входят сложные эфиры салициловой кислоты, метиловый эфир антраноловой кислоты, гелиотропин, линалоол, терпинеол, а также флавоноиды: робинин. Кора содержит ядовитый токсальбумин робин.
Заготовка сырья, первичная обработка и сушка. Цветки заготавливают во время цветения и сушат на чердаках или под навесами при хорошей вентиляции, разложив их в один слой на бумаге или ткани, или в сушилках при температуре 55-60°С.
Цветки заготавливают во время цветения и сушат на чердаках или под навесами при хорошей вентиляции, разложив их в один слой на бумаге или ткани, или в сушилках при температуре 55-60°С.
Стандартизация. Качество сырья регламентируется требованиями ВФС 42-1284-83.
Внешние признаки. Сырье состоит из соцветий, отдельных цветков и их частей. Морфологическую характеристику см. выше. Венчик белый или желтоватый; чашечка, цветоножка и ось соцветий зеленой или буровато-зеленой окраски. Запах ароматный. Вкус сладковатый.
Микроскопия. При рассмотрении с поверхности наружного и внутреннего эпидермиса чашечки видны клетки с прямыми, редко со слабоизвилистыми стенками. Устьица только с наружной стороны, с 5-6 околоустьичными клетками (аномоцитный тип). На чашечке, цветоножке и оси соцветия видны многочисленные, простые, прямые или изогнутые волоски с тонкими стенками, состоящие из короткой базальной и длинной терминальной клетки, реже — одноклеточные.
Наружный эпидермис верхнего лепестка (флага) имеет округло-многоугольные клетки с прямыми или извилистыми стенками и радиальными извилистыми складками кутикулы; внутренний эпидермис состоит из шестиугольных клеток с прямыми или извилистыми стенками и продольной складчатостью кутикулы. Эпидермис крыльев и лодочки отличается более выраженной складчатостью кутикулы. Вдоль жилок чашелистиков и лепестков проходят млечники с желтовато-оранжевым содержимым. С наружной стороны лепестков изредка встречаются простые одноклеточные волоски.
Числовые показатели.
Содержание робинина не менее 1,5%; потеря в массе при высушивании не более 11%; золы общей не более 7%; почерневших цветков не более 3%; других частей акации (листья, плоды, веточки) не более 7%; органической примеси не более 2%; минеральной примеси не более 1%.Хранение. Хранят в хорошо проветриваемых помещениях. Срок годности 3 года.
Использование. В народной медицине цветки применяются как жаропонижающее и спазмолитическое средство. Гипоазотемическое средство, продукт для получения фларонина.
Гипоазотемическое средство, продукт для получения фларонина.
28.06.2015
Акация серебристая: описание и фото
Изысканное австралийское дерево ― акация серебристая уже давно стал одним из самых распространенных растений на черноморском побережье России. У нас акацию часто называют мимозой. Ее пушистые, густо обсыпанные мелкими желтыми цветочками, ветки являются традиционным предвестником весны. На самом деле, это совершенно разные растения, хотя и принадлежащие одному семейству ― бобовые. Впрочем, на популярности акации эта путаница не сказывается: завораживающее впечатление от буйно цветущей пышной и плотной солнечной цветочной шапки никого не оставляет равнодушным.
Акация серебристая: ботаническая характеристика
Акация серебристая (Acacia dealbata) или акация подбеленная, принадлежит к роду Акация, семейству Бобовые. На родине, в юго-восточной части австралийского континента и на острове Тасмания, оно представляет собой раскидистое вечнозеленое дерево, высота которого может достигать 45 метров. Встречается в эвкалиптовых лесах, вдоль речных берегов. В Европе акация серебристая появилась в 1820 году, после чего распространилась на средиземноморском побережье. В России культура известна с 1852 года. Название «серебристая» растение получила по окрасу листвы. Ее оттенок из-за опушения листьев кажется пепельно-зеленым.
Встречается в эвкалиптовых лесах, вдоль речных берегов. В Европе акация серебристая появилась в 1820 году, после чего распространилась на средиземноморском побережье. В России культура известна с 1852 года. Название «серебристая» растение получила по окрасу листвы. Ее оттенок из-за опушения листьев кажется пепельно-зеленым.
В России в обиходе акацию серебристую часто неправильно называют мимозой. Серебристая акация широко распространена на Черноморском побережье Кавказа, и именно ее цветущие побеги являлись традиционным подарком в бывшем СССР в Международный женский день.
Акация серебристая – вечнозеленое дерево, имеющее густую зонтиковидную крону. Его высота в среднем равна 10-12 метрам (на родине в Австралии до 45 м). Обладает мощной корневой системой, расположенной горизонтально. Главный корень дает большое количество отпрысков.
Диаметр ствола может достигать 60-70 см. Покрывающая его кора гладкая, меняет цвет в зависимости от возраста: чем старше дерево, тем более темным становится оттенок.

Камедь – застывшие на стволе или ветках дерева капли густой жидкости. Она имеет свойство загущать (попадая в воду, набухает и образует клейкий раствор). Используется в различных областях промышленности, в том числе в медицине.
Листья по форме двояко-перистые, рассеченные на тонкие доли, длиной 10-20 см, покрытые множеством мелких волосков, состоящие из 8-24 пар длинных и узких мелких листочков первого порядка. В свою очередь каждый из них имеет до 50 пар еще более мелких и узких листочков второго порядка. Предназначение волосков – защита поверхности листьев от потери влаги, солнечных ожогов и холода. У оснований листиков первого порядка на тыльной поверхности главной жилки, находятся круглые железки. Когда акация цветет, из них сочится медовая жидкость.
Цветки серебристой акации представляют собой небольшие головки-бусинки серовато-желтого цвета, диаметром 4-8 мм. Эти мелкие шарики собраны в пышные кистевидные соцветия, а те в свою очередь собраны в соцветия-метелки. Лепестки, составляющие цветки яйцеобразные или широколанцетные, в венчике по 5 лепестков. Тычинки располагаются на длинных нитях и далеко выступают из венчика. Окрас пыльников ярко-желтый, а тычинок близкий к оранжевому. Цветение акации серебристой в зависимости от климатических условий приходится на период с середины зимы до середины весны.
Плоды акации серебристой ― продолговатые уплощенные с тупыми кончиками бобы длиной 1,5-8 см и шириной около 1 см, светло-коричневого или коричневого с фиолетовым отливом цвета. Внутри плодов располагаются твердые черные или темно-коричневые эллиптические семена длиной 3-4 мм. Плодоношение приходится на август и сентябрь.
Рост у серебристой акции быстрый. Сеянцы первого года жизни достигают высоты 50 см, а уже на второй год вырастают до 2,5 м. Некоторые из них в это время дают первое цветение. Обильное появление цветов и плодоношение начинается у растений, возраст которых равен 3-4 годам. Взрослые акации нередко ломаются от снега и сильных порывов ветра.
Некоторые из них в это время дают первое цветение. Обильное появление цветов и плодоношение начинается у растений, возраст которых равен 3-4 годам. Взрослые акации нередко ломаются от снега и сильных порывов ветра.
Из-за активного роста и неприхотливости к условиям произрастания эту разновидность нередко применяют для облесения склонов, лишенных иной растительности. К тому же акация серебристая принадлежит к числу ценных технических растений. Ее древесину используют в качестве топлива, для изготовления предметов народных промыслов. А получаемые из коры дерева дубильные экстракты применяют в кожевенном производстве.
Акация серебристая и мимоза: отличия
Внешний вид цветущих ветвей мимозы хорошо всем известен. Ее золотисто-желтоватые, солнечные цветки давно считаются символами наступающей весны. Но биологи знают, что на самом деле акация и мимоза совершенно разные растения. Деревце с пушистыми «цыплячьими» веточками, которые давно уже стали для нас символом весны, в действительности называется акацией серебристой и как по внешним, так и ботаническим характеристикам сильно отличается от мимозы.
Разница в том, что настоящие мимозы, в отличие от акации серебристой, произрастают в тропическом климате. В наших широтах мимозу можно вырастить только как однолетник и в тепличных условиях. Единственные места, где ее можно встретить в России ― оранжерейные коллекции. В зимний период она гибнет из-за дефицита тепла и света. Встретить ее в России можно только в оранжерейных коллекциях. А самой известной из более чем 300 видов мимоз считается мимоза стыдливая (Mimosa pudica) ― невысокий тропический полукустарник. Интересно растение тем, что его листья способны при прикосновении складываться попарно, а затем и поникать.
Как вырастить акацию серебристую в домашних условиях
Акация серебристая любит свет и тепло и отличается низкой морозостойкостью. Она способна расти только в благоприятных климатических, тропических или близких к ним условиях. В средней полосе это великолепное деревце культивируют как комнатное растение. При должном уходе акклиматизация акации серебристой в домашних условиях квартиры или загородного дома проходит прекрасно, а свидетельствует об этом ее обильное и продолжительное цветение.
История сотворения мира в древнеегипетской мифологии начинается с рассказа о том, как из вод изначального океана Нуна появился первозданный холм. И на холме этом выросла священная нильская акация-мимоза — «рука Атума». Мимоза — дочь Атума-Солнца, и потому ее соцветия состоят из маленьких желтых шариков-солнышек.
Размножение семенами
Выращивание из семян в домашних условиях лучше начинать проводить в январе. Для их активного прорастания нужно сделать легкий рыхлый грунт из одинаковых частей речного песка и листовой земли, добавив в смесь вермикулит. Семена также необходимо подготовить: замочить на пару суток в отстоянной воде, разогретой до 60 ºС, добавив в нее 1-2 капельки какого-либо биостимулятора роста. Во время замачивания нужно следить, чтобы вода с семенами не закисла.
Подготовленные семена помещают в контейнер с субстратом с заглублением 0,5 см, хорошенько поливают и накрывают прозрачным полиэтиленом или стеклом. Затем контейнер с семенами ставят в теплое и хорошо освещаемое место. Температура в помещении должна постоянно поддерживаться в пределах 22-25 ºС. Сроки прорастания семян в зависимости от их качества 2-5 недель.
Температура в помещении должна постоянно поддерживаться в пределах 22-25 ºС. Сроки прорастания семян в зависимости от их качества 2-5 недель.
Когда на саженцах акации серебристой появляются по 3-4 листика, их по отдельности пересаживают в горшочки с субстратом из смеси речного песка, перегноя, дерновой и листовой земли в соотношении 1:1:2:4.
Поскольку акация комфортно себя чувствует только при хорошем освещении, ей отводят комнату с окнами на юг. В конце осени и зимой в условиях короткого светового дня акацию обеспечивают дополнительной подсветкой в течение 3-4 часов. Если все делается правильно, саженцы быстро вырастают в молодые деревца и уже на второй год после посадки начинают цвести.
Выращивание из черенков
Черенкование акации серебристой в домашних условиях проводят весной, когда дерево отцветет, или в конце лета. Для этого отлично подходят побеги и ветки, оставшиеся после обрезки. Длина черенков должна быть около 10 см. Перед высадкой их 6-8 часов выдерживают в растворе Циркона или другого стимулятора роста.
Для укоренения очень удобно использовать специальные мини-теплички. Если их нет, подойдут обычные отдельные контейнеры, которые накрывают полиэтиленом или стеклянными банками, создав таким образом влажный и теплый микроклимат, необходимый черенкам для успешного укоренения и последующего роста. Грунт используют такой же, как описан выше.
Емкости или мини-теплички с саженцами ставят на солнечный теплый подоконник. Температура в помещении должна постоянно поддерживаться в диапазоне 22-24 ºС. После посадки черенки регулярно поливают, с промежутками в 3-5 дней. Зимой полив сокращают, до 1 раза в 10 суток. Весной и летом проводят комплексные подкормки. Если все сделано правильно, черенки укореняются спустя 2-3 месяца.
Цветки акации содержат эфирное масло, которое используется в парфюмерной промышленности. Кора акации серебристой также является ценным продуктом, она широко применяется в народной медицине.
Особенности ухода
Уход за серебристой акацией сводится к подкормкам, пересадкам, обрезке и поддержке грунта в состоянии умеренной влажности.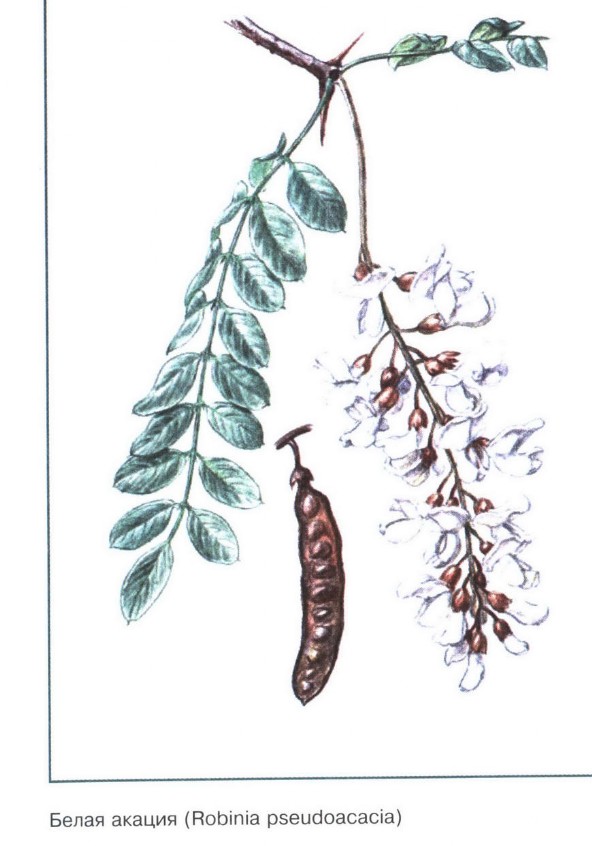
В период вегетации растение один раз в 3 недели подкармливают раствором какого-либо комплексного удобрения. Когда акация отдыхает, подкормки прекращают.
Растет акация быстро, переваливать ее приходится ежегодно. Растение обильно поливают, осторожно вытаскивают из старого горшка вместе с земляным комом, закрывающим корневую систему, устанавливают в центр емкости большего размера и добавляют вокруг ствола недостающий грунт.
В период вегетации акацию поливают до 2 раз в неделю, в период покоя ― 1 раз в 10 дней. В жару частоту поливов увеличивают и по необходимости растение опрыскивают.
Поскольку акация серебристая растет очень активно, ее нужно время от времени обрезать. Если этого не делать, крона загущается, растение может подвергаться различным заболеваниям. Обрезка выполняется после завершения цветения.
Летом деревце желательно перенести на лоджию, ему нужен свежий воздух. Если растение культивируется на даче или в загородном доме в теплое время года можно прикопать его в саду вместе с горшком, в котором оно растет, выбрав солнечный, хорошо освещенный участок.
При выращивании южной «гостьи» в европейской части России, например в Подмосковье, необходимо учитывать ряд особенностей. В первую очередь опытные садоводы рекомендуют оберегать акацию от существенных понижений или повышений температуры.
Болезни серебристой акации
Выращивая акацию серебристую, садоводы зачастую сталкиваются с рядом проблем:
- Акация сбрасывает листья, осыпаются бутоны. Этот процесс вызывается пересыханием грунта.
- Листья увядают. Причиной служит чрезмерный полив или, напротив, слишком сухая почва, а также очень тяжелый субстрат.
- Потеря яркого окраса листвы, которая становится бледной, возникает при недостатке света.
- Кончики листьев могут засыхать. Это свидетельствует о недостаточной увлажненности почвы и воздуха.
- Листва покрывается темными пятнами. Такое явление становится следствием воздействия сквозняков и холода.
- Появление паутинного клеща. Эта неприятность нередко бывает связано с низким уровнем влажности в помещении.

Для того чтобы избежать перечисленных проблем, необходимо тщательно следить за условиями содержания растения. При необходимости следует регулировать полив, уровень освещенности, влажность воздуха и почвы.
Все статьи об акации на сайте можно прочитать, пройдя по этой ссылке…
Акация серебристая нашла широкое применение в самых разных областях жизни. Это красивоцветущее солнечное дерево не слишком прихотливо в выращивании, а позитивные эмоции, которыми оно наполняет атмосферу дома, поистине бесценны.
Белая акация — описание, характеристики и советы по применению быстрорастущего дерева в ландшафтном дизайне сада (фото + видео)
А что, если я вам скажу, что белая акация, которую мы видели с самого детства, на самом деле акацией не является? Многие подумают, что это бред, но ботаники называют это растение иначе, и среди его названий встречаются такие, как робиния и робиния лжеакациевая. О том, что это за дерево и о многих других интересных вещах мы сегодня и поговорим.
Краткое содержимое статьи:
История
Если не все, то очень многие должны представлять, как выглядит акация обыкновенная. Несмотря на это название считается не совсем корректным.
Робинией это дерево назвал Карл Линней, который тем самым отдал должное своим французским ботаническим коллегам – братьям Веспасиану и Жану Робэн. Это особенно забавно, если вспомнить, что все понимают, что такое акация, а услышав название робиния, быстрее вспомнят о рябине, но уж никак не о том, о чем думал Линней.
В ботаническом плане только одно название является подлинным, но будьте свободны использовать то, какое вам удобнее.
Описание
Впервые дерево акации было обнаружено в лесах Северной Америки. Несмотря на то, что акация предпочитает более теплый, южный, климат, она отлично акклиматизировалась почти во всех местах, куда ее завезли. Исключением стали только места нашей планеты, где лежит снег практически круглый год.
На территории стран СНГ акация является очень популярным деревом, что во многом заслуга ее лечебных свойств, о которых мы поговорим позже.
Коренные индейцы называли акацию деревом жизни и источником бессмертия. Древние племена верили, что ее цветки являются символом чистоты и процветания.
Цветет дерево в первые летние месяцы, и в это время активнее всего приходится работать пчелам, которые собирают прекрасный мед из цветков этого замечательного дерева.
Ученым удалось подсчитать, что с одного выросшего дерева пчелы могут собрать от пяти до пятнадцати килограмм качественного меда, который хорош не только своими полезными свойствами, но и не кристаллизуется, что определенно является плюсом. Добавьте к этому еще и то, что мед является очень вкусным, и вы получите дерево, которое должен иметь каждый пасечник на своем участке.
Независимо от вида акация растет достаточно быстро, и уже в первые десять лет может добраться до отметки в тридцать метров в высоту, что по сути очень сильный показатель. Дерево отлично справляется даже с не очень благоприятными условиями, такими, как малое количество влаги, и морозы зимой. Поэтому его стали часто использовать при создании лесополос для защиты от ветра. Корневая система очень большая и крепка, из-за чего акацию часто используют для укрепления почвы с высоким содержанием песка.
Дерево отлично справляется даже с не очень благоприятными условиями, такими, как малое количество влаги, и морозы зимой. Поэтому его стали часто использовать при создании лесополос для защиты от ветра. Корневая система очень большая и крепка, из-за чего акацию часто используют для укрепления почвы с высоким содержанием песка.
Теперь, если отбросить функционал, так сказать, и вспомнить о внешнем виде, то можно заметить, что это дерево отлично подходит и для ландшафтного дизайна, ведь высокое растение с пышной и красивой короной может украсть любой участок.
Интересный факт
То, что белая акация дерево очень интересное и особенное заметил и Генрих VI, который в далеком семнадцатом веке распорядился, чтоб его посадили в королевском саду в Париже. Сейчас белая акация является самым старым деревом в городе любви, а ведь прошло уже более четырех сотен лет.
Удивительно, но за это время очень много изменилось, и сейчас акацию можно встретить даже в миниатюрной форме, которую вывели японские ботаники.
Акация, выращенная как бонсай, является очень популярной во многих странах мира, ведь такое растение можно держать даже в небольшой квартире, ну и конечно же перевозить с собой без проблем.
Лечебные свойства
Независимо от того, белая или розовая акация, плоды дерева обладают прекрасными лечебными свойствами. Из цветков можно делать настойки, сиропы и многие другие средства. Они могут помочь от множества проблем со здоровьем, ведь помогают справиться со спазмами, понизить жар, способствуют ускорению свертываемости крови и заживлению ран, решить проблему непроходимости кишечника и многие другие полезные свойства.
Если вам приходилось видеть, как цветет акация, то вы можете представить тот замечательный запах, который издают ее цветки. Мед из белой акации и лечебные средства — это еще более интересная история.
То, что сейчас известно, как народные средства, сотни лет назад было аналогом медицины. Все возможные проблемы старались вылечить при помощи природных средств, и акация была одним из них.
Все возможные проблемы старались вылечить при помощи природных средств, и акация была одним из них.
Сейчас уже доказано, что она может быть использована в устранении воспаления и опухолей, профилактике ревматических заболеваний и проблем с суставами, простуды и даже переутомлением и депрессией, которая следует за этим.
Стоит отметить еще и то, что ученые открыто признали, что даже сейчас они не полностью изучили все свойства акации, или же как они ее называют – робинии, а потому еще не уверенны в пределах, в которых она может быть использована.
В любом случае это прекрасное и полезное дерево, которое отлично подходит для нашего климата и может стать источником меда и лекарственных средств.
Фото белой акации
Также рекомендуем просмотреть:
Белая акация. Уход, размножение, выращивание. Фото — Ботаничка.ru
Воздух южных городов и сел в пору цветения белой акации напоен ее ароматом, знаменующим приход щедрой летней поры. Это дерево воспето в старинных романсах, во многих песнях, не обошли его вниманием и в современных художественных произведениях. Аромат акации далеко разносится ветром по полям. Нектар ее неодолимо влечет пчел. В цветущем акациевом насаждении на площади в один гектар они собирают более 1500 килограммов меда, а со среднего по размерам дерева могут собрать около 8 килограммов.
Аромат акации далеко разносится ветром по полям. Нектар ее неодолимо влечет пчел. В цветущем акациевом насаждении на площади в один гектар они собирают более 1500 килограммов меда, а со среднего по размерам дерева могут собрать около 8 килограммов.
Свежий мед из белой акации отличается отменным вкусом, целебными свойствами, тонким запахом. Он почти бесцветен и удивительно прозрачен — в сотах или доверху налитом стеклянном сосуде его не всегда можно заметить. Акациевый мед долго сохраняет жидкое состояние, а уж если и кристаллизуется, то не теряет пищевых качеств.
Белая акация — наиболее распространенное дерево юга нашей страны. Безраздельно господствует она в степной части Украины, на Кубани, в Молдавии. Нельзя представить Кишинев и Одессу, Днепропетровск и Ростов, Ворошиловград, Донецк, Краснодар и многие другие города нашего юга без белой акации. Но самое удивительное то, что каких-нибудь 200 лет назад ее здесь вовсе не было.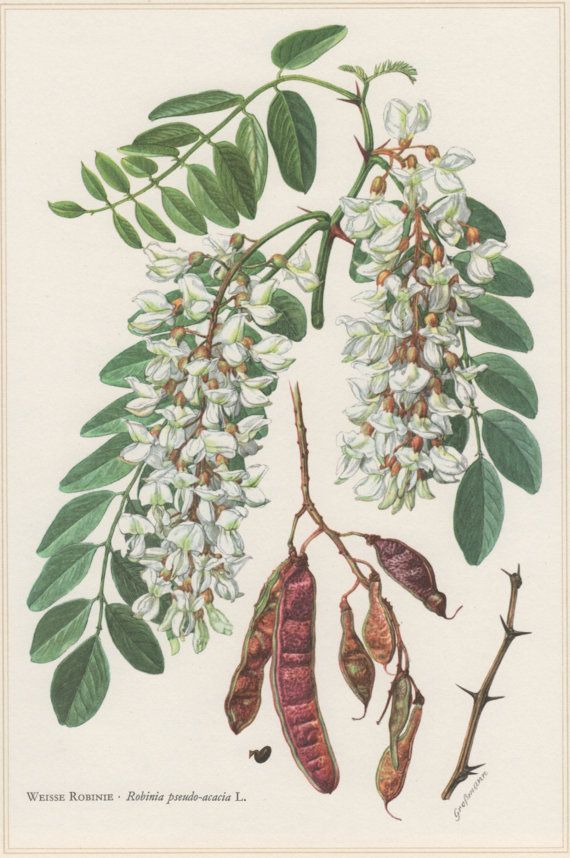 Теперь только специалисты знают, что белая акация завезена к нам из Северной Америки, где она растет в обширных естественных лесах.
Теперь только специалисты знают, что белая акация завезена к нам из Северной Америки, где она растет в обширных естественных лесах.
По утверждению ботаников, акация была одним из первых деревьев, привезенных из Нового Света в Европу. Вывез ее из штата Виргиния путешествовавший по Америке садовод Людовика XIII Веспасиан Робин.
Карл Линней, разработавший в первой половине XVIII века систему классификации растительного мира, дал роду, к которому отнесена белая акация, в честь Робина научное латинское название робиния. Позднее ботаники стали называть белую акацию также ложной акацией в отличие от многочисленных видов рода настоящих акаций, распространенных преимущественно в тропических странах.
Белая акация, или Робиния псевдоакация, или Робиния лжеакация, Робиния обыкновенная (Robinia pseudoacacia). © SiebrandПервое дерево, которое посадил сам Робин в 1635 году в Париже в Ботаническом саду французской академии наук, сохранилось как своего рода исторический памятник до наших дней. Теперь белая акация широко распространилась не только у нас в стране, но и растет на всех континентах Земли, исключая Антарктиду. Ни одна порода, не считая, может быть, нашу березу, не может сравниться с ней по способности быстро обживать новые территории. Правда, «метод» освоения новых мест у нее свой: береза щедро рассевает семена, а акация завоевывает жизненное пространство корневыми отпрысками.
Теперь белая акация широко распространилась не только у нас в стране, но и растет на всех континентах Земли, исключая Антарктиду. Ни одна порода, не считая, может быть, нашу березу, не может сравниться с ней по способности быстро обживать новые территории. Правда, «метод» освоения новых мест у нее свой: береза щедро рассевает семена, а акация завоевывает жизненное пространство корневыми отпрысками.
Белая акация не на последнем месте и по семенной производительности — она дает весьма обильные урожаи семян. Лесоводы утверждают, что из семян, собранных лишь в течение одного года со среднего по величине и возрасту дерева, можно вырастить более 200 тысяч сеянцев белой акации. Тем не менее, в природных условиях белая акация семенным путем почти не возобновляется, уж очень твердая и плотная у ее семян оболочка. Поэтому лесоводы перед посевом несколько раз обваривают ее семена кипятком.
Белая акация, или Робиния псевдоакация, или Робиния лжеакация, Робиния обыкновенная (Robinia pseudoacacia). © H. Zell
© H. ZellУ нас белую акацию впервые высадили в начале XIX века в саду А. К. Разумовского под Одессой, откуда она вскорости была позаимствована Одесским ботаническим садом. Примерно тогда же семена белой акации выписал непосредственно из Северной Америки Василий Назарович Каразин, основатель Харьковского университета. В Одессе, Киеве и на Харьковщине растут самые старые в нашей стране акации, чей возраст значительно превышает 100 лет, а размеры их поражают даже знатоков. Одно из таких деревьев-старожилов растет в Ботаническом саду Киевского университета.
Сохранились на Украине и памятные деревья этой экзотической породы. Одно из них особенно дорого почитателям великого кобзаря — Тараса Шевченко. В Переяславе-Хмельницком возле домика большого друга поэта, врача Козачковского растут две старые акации, стволы которых тесно переплелись. В свое время Шевченко и Козачковский высадили два саженца акации в одну ямку, а стволики накрепко сплели. Сохранилось предание, что, закончив посадку, Шевченко крепко пожал руку Козачковскому и сказал: «Пусть и люди русские и украинские братаются, подобно нашим деревьям»
Белая акация, или Робиния псевдоакация, или Робиния лжеакация, Робиния обыкновенная (Robinia pseudoacacia). © Doronenko
© DoronenkoИспользуемые материалы:
- С. И. Ивченко — Книга о деревьях
Акация белая лекарственное растение
Семейство Бобовые – Fabaceae.
Народные названия: робиния ложно-акациевая, гороховое дерево, караган.
Ботаническое описание. Дерево семейства бобовых (Fabaceae), до 25-30 м (в среднем 12 м) высоты, с раскидистой кроной,
толстой, темной, глубокобороздчатой корой. Акация белая имеет очень
мощную корневую систему, корни которой проникают на глубину до 20 и более метров. Листья
непарноперистые, на коротких черешках, которые в направлении к ветви
видоизменяются в хрящевидные ложные влагалища. Лист составляет от 4 до 8
пар эллиптических, светло-зеленых, мягких, цельнокрайних листочков. Шипы
(видоизмененные листья) – крепкие, деревянистые, с очень острым концом,
серповидные. Цветки многочисленные, ароматные, белые, крупные, в
повислых кистях. Плод – плоский боб до 10 см длины, с семенами темной
окраски. Цветет в мае – июне, плоды созревают в сентябре и остаются
висеть на дереве всю зиму. Продолжительность жизни – до 50 лет и более.
Растет вдоль дорог, в садах и парках. Растение ядовито (семена, кора,
корни)! Родина – Северная Америка, где встречается в северо-восточных и
северных штатах от Пенсильвании до Дакоты, Айовы, Монтаны и Оклахомы. В
1804 году семена этого растения были впервые высажены в России, в 1813
году оно появилось в Крыму в Никитском ботаническом саду.
Цветет в мае – июне, плоды созревают в сентябре и остаются
висеть на дереве всю зиму. Продолжительность жизни – до 50 лет и более.
Растет вдоль дорог, в садах и парках. Растение ядовито (семена, кора,
корни)! Родина – Северная Америка, где встречается в северо-восточных и
северных штатах от Пенсильвании до Дакоты, Айовы, Монтаны и Оклахомы. В
1804 году семена этого растения были впервые высажены в России, в 1813
году оно появилось в Крыму в Никитском ботаническом саду.
Сбор и сушка сырья. Лекарственным сырьем являются цветки акации белой, которые собирают в начале цветения в полураспустившемся состоянии. Сушат в тени под навесом, в хорошо проветриваемом помещении или в сушилках при температуре 40-50°С, рассыпая тонким слоем и часто переворачивая. Кору и листья собирают в течение всего периода вегетации.
Действующие вещества. В цветках и коре растения содержится робинин. Наиболее высокое содержание
робинина приходится на период, когда бутоны набухают, но еще не распустились. Когда цветки распускаются, содержание его понижается. В цветках также обнаружены
акацин, бикробин, бикверцитин, эфирное масло. Эфирное масло содержит метиловый
эфир антраниловой кислоты, индол, гелиотропин, линалоол, терпинеол. Представляет
собой полужидкую массу светло-желтого цвета с приятным запахом цветков акации. В
древесине содержатся флавоноиды робинетин, дигидроробинетин, физетин, робтин,
робтеин, бутеин, ликвартигенин и др. В листьях найдены гликозиды индикан и
сирингин, витамин А, аскорбиновая кислота, флавоноиды (акацетин и его гликозид
акациин, апигенин, кверцетин, лютеолин) и дубильные вещества.
Когда цветки распускаются, содержание его понижается. В цветках также обнаружены
акацин, бикробин, бикверцитин, эфирное масло. Эфирное масло содержит метиловый
эфир антраниловой кислоты, индол, гелиотропин, линалоол, терпинеол. Представляет
собой полужидкую массу светло-желтого цвета с приятным запахом цветков акации. В
древесине содержатся флавоноиды робинетин, дигидроробинетин, физетин, робтин,
робтеин, бутеин, ликвартигенин и др. В листьях найдены гликозиды индикан и
сирингин, витамин А, аскорбиновая кислота, флавоноиды (акацетин и его гликозид
акациин, апигенин, кверцетин, лютеолин) и дубильные вещества.
Применение в медицине. Кора (молодых деревьев). В
народной медицине отвар – при обострении гастритов и язвенной болезни. В
Болгарии и Германии спиртовая настойка – при повышенной кислотности желудочного
сока и других заболеваниях желудка. Такое же применение имеет в гомеопатии.
Листья, молодые побеги. В народной медицине настойка – при обострении гастритов
и язвенной болезни желудка и двенадцатиперстной кишки. Цветки. В народной
медицине настой, отвар – при заболеваниях почек и мочевого пузыря. Настойка
наружно – при ревматизме. Цветки акации белой используются в смеси с листьями
толокнянки обыкновенной, цветками пижмы обыкновенной, цветками василька синего,
корнем солодки (как мочегонный сбор). В Болгарии настой – как отхаркивающее и
жаропонижающее средство.
В народной медицине настойка – при обострении гастритов
и язвенной болезни желудка и двенадцатиперстной кишки. Цветки. В народной
медицине настой, отвар – при заболеваниях почек и мочевого пузыря. Настойка
наружно – при ревматизме. Цветки акации белой используются в смеси с листьями
толокнянки обыкновенной, цветками пижмы обыкновенной, цветками василька синего,
корнем солодки (как мочегонный сбор). В Болгарии настой – как отхаркивающее и
жаропонижающее средство.
Лекарственные формы, способ применения и дозы.
Отвар коры акации: 1/2 столовой ложки измельченного сырья заливают 500 мл воды, кипятят 20 мин, процеживают горячим и доводят объем кипяченой водой до исходного. Принимают небольшими порциями в течение 2 дней. При хорошей переносимости можно выпивать в течение дня, лучше в теплом виде.
Настойка листьев и молодых побегов акации: сырье настаивают на 40% спирте в соотношении 1:10, выдерживая 15 дней, периодически взбалтывая.
Принимают по 1 чайной ложке 3 раза в день до еды.
Настой цветков акации: 10 г сырья заливают 200 мл кипятка, настаивают, затем процеживают.
Отвар цветков акации: 1 столовую ложку свежего или сушеного сырья заливают 500 мл воды, кипятят 3 мин, процеживают и доводят объем до исходного.
Принимают по 1 столовой ложке 3-4 раза в день до еды.
Настойка цветков акации: сырье заливают спиртом или водкой и настаивают в течение 2 недель на солнце. Применяют наружно.
Побочные эффекты. При внутреннем применении лекарственных форм (семена, кора, корни) следует соблюдать
осторожность ввиду ядовитости. Острые отравления, которые сопровождаются
недомоганием, тошнотой, рвотой, головной болью или сонливостью, могут возникнуть
также при заготовке древесины.
К алкалоидам акации особенно чувствительны лошади. При отравлении у животных появляются сильные боли в
животе. У них отмечаются сильное возбуждение, дрожь, судороги, слабость
сердечной деятельности. Первая помощь заключается в промывании желудка,
назначении адсорбирующих средств, применении симптоматического лечения
(возбуждающие, сердечные и другие средства).
Первая помощь заключается в промывании желудка,
назначении адсорбирующих средств, применении симптоматического лечения
(возбуждающие, сердечные и другие средства).
Акация белая | Дары природы.су
Автор Дары природы.су На чтение 2 мин. Просмотров 35 Опубликовано
Акация белая (Acacia dealhata) — это крупное дерево с яйцевидно-продолговатыми листьями. Цветки душистые, белые, собраны в поникающие кисти. Цветет в мае—июне.
В качестве лекарственного сырья используют кору молодых ветвей и нераспустившиеся цветки. Кору надо собирать осенью, а цветки — весной.
Настой цветков акации обладает жаропонижающим, противовоспалительным, отхаркивающим действием. Кору лучше использовать при расстройствах желудка и повышенной кислотности.
■ ВНИМАНИЕ!
В больших дозах это растение ядовито.Содержащийся в коре токсальбумин робин может вызвать раздражение слизистых оболочек. При отравлении отмечается нарастающее возбуждение.
Энергетические свойства. Акация отдает энергию, наполняя человека бодростью. Особенно полезна женщинам.
ГАСТРИТ С ПОВЫШЕННОЙ КИСЛОТНОСТЬЮ
Залить 1,5 чайной ложки измельченной коры акации 2 стаканами кипятка, настаивать в течение 2 часов. Принимать по 1 столовой ложке 3—-4 раза в день.
ГРИПП
Взять в равных частях сухие цветки акации белой и бузины черной.
Залить 4 столовые ложки смеси 3 стаканами кипятка, настаивать, укутав, 1 час, процедить. Выпить в течение дня, делая по нескольку глотков.
КОКЛЮШ
Залить 1 чайную ложку цветков акации 1 стаканом горячего молока, поставить на медленный огонь и держать 5—10 минут, добавить 1 чайную ложку меда. Пить как чай перед едой.
ЛИХОРАДКА, ВЫСОКАЯ ТЕМПЕРАТУРА
Залить 2 чайные ложки цветков акации 1/2 стакана водки, настаивать 1 неделю в закрытой посуде, процедить. Принимать по 20 капель, разбавив водой.
Принимать по 20 капель, разбавив водой.
ЦИСТИТ
Залить 1 чайную ложку цветков акации 1 стаканом кипятка, настаивать под крышкой 30 минут. Принимать по 1 столовой ложке 3 раза в день.
ПОДАГРА, АРТРИТ, АРТРОЗ, ТРОМБОФЛЕБИТ
Залить 30—40 г цветков 1 стаканом водки и настаивать в течение 7 дней.
Смазывать больные места 2—3 раза в день. Лекарство хорошо рассасывает вздувшиеся варикозные узлы.
ШПОРА ПЯТОЧНАЯ
Залить цветки белой акации водкой в соотношении 1:3. Настоять и смазывать шпоры.
Использованная литература: Травник: золотые рецепты народной медицины/ А. Маркова. — М.: Эксмо; Форум, 2007
Мне нравитсяНе нравитсяWhitethorn Acacia — DesertUSA
Акация констрикта
Кустарник белой акации рядом с небольшим арройо, раскидистые многоствольные ветви.
Чаще всего акация боярышник — бобовое или бобовое растение,
растение — представляет собой прямостоячий, древесный, полувечнозеленый куст с
несколько стволов. Обычно он вырастает от нескольких футов до 15 или 16 футов в высоту.
часто образует густые заросли.
Обычно он вырастает от нескольких футов до 15 или 16 футов в высоту.
часто образует густые заросли.
Отдельное растение может выглядеть почти пенистым, с мелким зеленоватым или сероватым оттенком. кружевные листья.Часто теряет листья в холодную погоду или длительное время. засуха. Цветет весной, а иногда и в конце лета. миниатюрный желтый шар цветет и источает сладкий ночной аромат пустыни. Во всем мире насчитывается около 800 видов.
Отличительные особенности
Стволы, ветки и веточки: Многоствольные и ветвистые со слегка зигзагообразной формой веточки, более молодые, имеющие несколько красноватый цвет, и более старые приросты, серовато-коричневый.
Кора: Довольно гладкая на молодых ветвях и веточках, несколько грубая и грубая. бороздчатые или бороздчатые на старых стволах и ветвях.
Корни: Широкораспространенные боковые корни и длинный проникающий главный корень, достигающий
как для мелководья, так и для более глубоких вод (согласно Volney M. Spalding, Distribution
и движение пустынных растений ). В корнях обитают бактерии, которые могут усваивать азот
из воздуха и превращают его в соединения, обогащающие почву.
Spalding, Distribution
и движение пустынных растений ). В корнях обитают бактерии, которые могут усваивать азот
из воздуха и превращают его в соединения, обогащающие почву.
Шипы: Примерно 1/2 дюйма длиной, острая игла, парные и белый, что дало «белошовную акацию» ее общее название.Более очевидный на более молодых ветках и молодых растениях.
Листья: Несколько дюймов в длину, «двупородное соединение» или «дважды соединение », что означает, что лист состоит из нескольких пар основных листочков, каждая из которых, в свою очередь, состоит из нескольких пар мелких листочков.
Цветение: От 1/4 до 1/3 дюйма в диаметре, ярко-желтая, плотная сфера тычинок, которые предлагают насекомым минимальное количество пыльцы и отсутствие нектара. В результате цветет привлечь относительно немного опылителей.
Стручки фасоли: Тонкие, красновато-коричневые, капсулы длиной от двух до пяти дюймов с характерными
сужения между бобами, что придает Acacia constricta научный
имя.
Пернатые листья и спелые бобы Whitethorn Acacia
Местное распространение и среда обитания
Белая акация естественным образом растет на юге Аризоны, на юге Нью-Мексико. западный Техас и северная и центральная Мексика, населяющие берега рек, поймы, прерывистый дренаж, каменистые склоны и холмы на высотах от От 2000 до 6000 футов.
Регенерация
Белая акация прорастает из семян, но может прорастать из корня. кроны в случае повреждения или гибели ее стволов и ветвей, например, в результате пожара.
Опыление и производство семян: Растение опыляется пчелами и другие насекомые, которые часто посещают недолговечные цветы только в первый день они открывают. При отсутствии у насекомых достаточного интереса растение может самоопыляться, но его урожай фасоли, вероятно, значительно снизится.
Распространение и прорастание семян: Распространение семян белой акации
в первую очередь птицами, особенно перепелами, и млекопитающими, особенно грызунами. Семена, с которых была удалена оболочка при прохождении через птичий или
пищеварительный тракт млекопитающих прорастает легче. Некоторые семена кешируются
грызунами под поверхностью почвы, защищенными от лесных пожаров.
Семена, с которых была удалена оболочка при прохождении через птичий или
пищеварительный тракт млекопитающих прорастает легче. Некоторые семена кешируются
грызунами под поверхностью почвы, защищенными от лесных пожаров.
Посевной материал: Семена заглублены на полдюйма или дюйм ниже поверхности почвы имеют наибольшую вероятность стать всходами, особенно если они прорастают при высоких температурах с небольшим количеством влаги.Семена на поверхности могут прорасти, но из них редко появляются всходы.
Созревание: Рост регулируется наличием влаги. белая акация обычно медленно созревает в пустынной среде. После создания он может жить семь десятилетий или больше.
Интересные факты
Акация белая принадлежит к одному из самых крупных семейств кустарников и деревья — бобовые, семейство гороховые. Он насчитывает около 400 родов и 10 000 видов.
Классический пример взаимовыгодных отношений природы: акации
в Центральной Америке и Южной Африке дают муравьям семена, которые несут питательные
жировое тело называется элайсоме или арил. Муравьи разносят семена в гнезда,
поедание элисомы и отбрасывание целого семени, но эффективная посадка
это для акации, способствуя ее распространению. От этого выигрывают и акация, и муравей.
Муравьи разносят семена в гнезда,
поедание элисомы и отбрасывание целого семени, но эффективная посадка
это для акации, способствуя ее распространению. От этого выигрывают и акация, и муравей.
Некоторые виды акации производят такую твердую и долговечную древесину, что ее использовали в качестве судостроительных гвоздей, вагонных осей и столбов ограждений.В отчетах говорится, что акация столбы забора прослужили иногда до века.
Индейцы используют
Доисторические американцы перемалывали бобы белой акации в пищу. в приготовлении пищи. Они использовали бобы, листья и корни для приготовления лекарственного чай, которым лечили желудочные и респираторные заболевания. Они использовали порошок бобы и листья, чтобы сделать припарку для лечения кожной сыпи. Они использовали цветы и листья для приготовления лечебного чая для снятия похмелья.
Современные жители пустыни сажают в садах акацию белошовную, зная, что
растение требует относительно мало воды, привлекает диких животных (например,
перепелов, голубей, пиррулоксий и других птиц) и обогащает окружающую почву.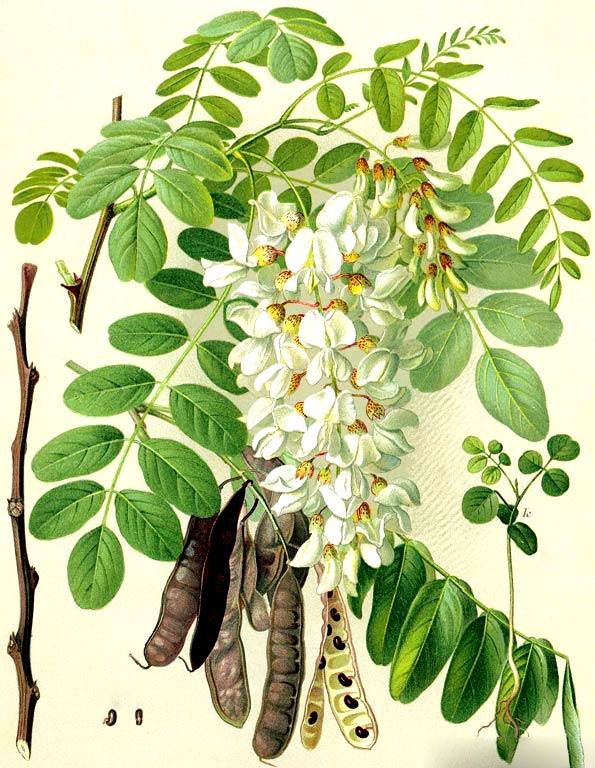
Джей У. Шарп
Связанные страницы DesertUSA
Поделиться этой страницей в Facebook:
Информационный бюллетень DesertUSA — Мы рассылаем статьи о походах, кемпингах и местах для изучения, а также о животных, сообщениях о полевых цветах, информации о растениях и многом другом.Зарегистрируйтесь ниже или , чтобы узнать больше о новостной рассылке DesertUSA здесь . (Это бесплатно.)
Окружающая среда пустыни
Пустыни Северной Америки
Геологические термины пустыни
Frontiers | Физиобиохимические и морфологические признаки галофитного бобового кустарника, проростков Acacia ampiceps в ответ на солевой стресс в теплице
Введение
Засоление почвы является одним из основных абиотических стрессов, серьезно влияющих на рост, развитие и продуктивность растений. Он поражает почти 397 миллионов гектаров (млн га) земли во всем мире, особенно в Азиатско-Тихоокеанском регионе и Австралии, где около 195 млн га засолены (Flowers et al., 1986; Rengasamy, 2006). Галофитные (солеустойчивые) виды, составляющие лишь 1% мировой флоры, развили солевой защитный механизм и демонстрируют оптимальный рост в засоленной почве (Flowers and Colmer, 2008). Такие механизмы солеустойчивости включают систему ионного гомеостаза через солевые железы / солевые пузыри, систему осморегуляции, такую как свободный пролин, глицинбетаин, маннит и растворимые сахара, гормональную регуляцию и антиоксидантный механизм (Hasegawa et al., 2000; Flowers et al., 2010; Шабала, 2013; Шабала и др., 2014). Гомеостаз Na + через антипортерные белки, т.е. обменник Na + / H + (NHXs) у галофитных видов, хорошо зарекомендовал себя в качестве основного канала для управления притоком Na + из почвенного раствора в корневую клетку. и перемещаются посредством нагрузки ксилемы в другие органы, что приводит к увеличению соотношения Na + / K + (Flowers and Colmer, 2008; Li et al.
Он поражает почти 397 миллионов гектаров (млн га) земли во всем мире, особенно в Азиатско-Тихоокеанском регионе и Австралии, где около 195 млн га засолены (Flowers et al., 1986; Rengasamy, 2006). Галофитные (солеустойчивые) виды, составляющие лишь 1% мировой флоры, развили солевой защитный механизм и демонстрируют оптимальный рост в засоленной почве (Flowers and Colmer, 2008). Такие механизмы солеустойчивости включают систему ионного гомеостаза через солевые железы / солевые пузыри, систему осморегуляции, такую как свободный пролин, глицинбетаин, маннит и растворимые сахара, гормональную регуляцию и антиоксидантный механизм (Hasegawa et al., 2000; Flowers et al., 2010; Шабала, 2013; Шабала и др., 2014). Гомеостаз Na + через антипортерные белки, т.е. обменник Na + / H + (NHXs) у галофитных видов, хорошо зарекомендовал себя в качестве основного канала для управления притоком Na + из почвенного раствора в корневую клетку. и перемещаются посредством нагрузки ксилемы в другие органы, что приводит к увеличению соотношения Na + / K + (Flowers and Colmer, 2008; Li et al. , 2008). С другой стороны, содержание свободного пролина и общего содержания растворимого сахара в галофитных растениях было зарегистрировано как основные осмолиты, когда растения подвергаются засолению почвы (Yokota, 2003; Patel et al., 2010; Theerawitaya et al., 2014). В случае физиологических изменений фотосинтетический пигмент, то есть хлорофиллы a и b, деградирует в зависимости от уровня содержания соли в почвенном растворе, особенно при экстремальном солевом стрессе (10 dS m –1 ) (Giri et al., 2003 ), что приводит к снижению чистой скорости фотосинтеза, особенно при сильном солевом стрессе (Takemura et al., 2000).
, 2008). С другой стороны, содержание свободного пролина и общего содержания растворимого сахара в галофитных растениях было зарегистрировано как основные осмолиты, когда растения подвергаются засолению почвы (Yokota, 2003; Patel et al., 2010; Theerawitaya et al., 2014). В случае физиологических изменений фотосинтетический пигмент, то есть хлорофиллы a и b, деградирует в зависимости от уровня содержания соли в почвенном растворе, особенно при экстремальном солевом стрессе (10 dS m –1 ) (Giri et al., 2003 ), что приводит к снижению чистой скорости фотосинтеза, особенно при сильном солевом стрессе (Takemura et al., 2000).
Галофиты считаются потенциально новыми культурами для использования в качестве кормовых, овощных и масличных культур. Однако потенциальное использование галофитных видов для роста в засоленных почвах и для облегчения фиторемедиации засоленных почв зависит от нескольких факторов, таких как накопление соли, относительная скорость роста и конверсия биомассы, многоцелевое использование и экономическая отдача для фермеров (Panta et al. ., 2014). Acacia ampiceps (акация), уроженец Австралии, представляет собой многоцелевой вид растений, широко используемый для кормления скота (из расчета 120–170 г на кг сырого протеина –1 ; Dynes and Schlink, 2002) и производства биомассы (23,34). % сухой биомассы; Ashraf et al., 2012). Кроме того, он имеет потенциал для восстановления засоленных почв и хорошо растет в почве с EC e ≥ 16 dS m –1 (Ansari et al., 2007). Ранее с использованием многомерного кластерного анализа продуктивности биомассы видов Acacia в засоленной почве, A.ampiceps , или ака, имеет максимальную продукцию биомассы (39,69 т / га –1 ), чем другие виды: A. karroo (2,39 т / га –1 ), A. stenophylla (0,53 тонна га –1 ), A. seyal (0,27 тонны га –1 ) и A. asak (0,18 тонны га –1 ) (Aref et al., 2003). В Таиланде для восстановления засоленных почв в засоленные районы на северо-востоке Таиланда был интродуцирован бобовый кустарник A.
., 2014). Acacia ampiceps (акация), уроженец Австралии, представляет собой многоцелевой вид растений, широко используемый для кормления скота (из расчета 120–170 г на кг сырого протеина –1 ; Dynes and Schlink, 2002) и производства биомассы (23,34). % сухой биомассы; Ashraf et al., 2012). Кроме того, он имеет потенциал для восстановления засоленных почв и хорошо растет в почве с EC e ≥ 16 dS m –1 (Ansari et al., 2007). Ранее с использованием многомерного кластерного анализа продуктивности биомассы видов Acacia в засоленной почве, A.ampiceps , или ака, имеет максимальную продукцию биомассы (39,69 т / га –1 ), чем другие виды: A. karroo (2,39 т / га –1 ), A. stenophylla (0,53 тонна га –1 ), A. seyal (0,27 тонны га –1 ) и A. asak (0,18 тонны га –1 ) (Aref et al., 2003). В Таиланде для восстановления засоленных почв в засоленные районы на северо-востоке Таиланда был интродуцирован бобовый кустарник A..jpg) ampiceps (Wongpokkhom et al., 2008). Однако защитные механизмы, лежащие в основе солевой толерантности в A. ampiceps , в значительной степени неизвестны. Однако исследований по изучению физиологических и биохимических механизмов, лежащих в основе механизмов солеустойчивости у A. ampiceps, не проводилось. Таким образом, мы провели серию экспериментов по изучению биохимических, физиологических и морфологических свойств A. ampiceps , выращенного в ответ на экстремальные концентрации соли в тепличных условиях.
ampiceps (Wongpokkhom et al., 2008). Однако защитные механизмы, лежащие в основе солевой толерантности в A. ampiceps , в значительной степени неизвестны. Однако исследований по изучению физиологических и биохимических механизмов, лежащих в основе механизмов солеустойчивости у A. ampiceps, не проводилось. Таким образом, мы провели серию экспериментов по изучению биохимических, физиологических и морфологических свойств A. ampiceps , выращенного в ответ на экстремальные концентрации соли в тепличных условиях.
Материалы и методы
Растительный материал и солевые обработки
Семена акации ( Acacia ampiceps Maslin), предоставленные Департаментом развития земель Министерства сельского хозяйства и кооперативов Таиланда, дезинфицировали 15% гипохлоритом натрия в течение 30 минут с последующей трехкратной промывкой водопроводной водой, а затем дистиллированной водой. Дезинфицированные семена проращивали в пластиковом лотке (30 × 60 × 5 см), содержащем 1 кг садовой почвы (супеси; ЕС = 2. 687 dS m –1 ; pH = 5,5; органическое вещество = 10,36%; общий азот = 0,17%; общий фосфор = 0,07%; общий калий = 1,19%) в тепличных условиях. 15-дневные проростки переносили в полиэтиленовый пакет (Ш × Д × В; 4 × 4 × 12 см), содержащий садовую почву (как указано выше), и выращивали в контролируемых условиях окружающей среды (относительная влажность 80 ± 5%, 500–1000 мкмоль м –2 с –1 плотность потока фотосинтетических фотонов, 10 ± 2 hd –1 фотопериод от солнечного света и температуры окружающей среды 28 ± 2 ° C) в течение 7 недель.Эксперимент проводился в оранжерее Тайского научного парка, Патхум Тани, Таиланд (14 ° 01′12 ″ северной широты и 100 ° 31′12 ″ восточной долготы).
687 dS m –1 ; pH = 5,5; органическое вещество = 10,36%; общий азот = 0,17%; общий фосфор = 0,07%; общий калий = 1,19%) в тепличных условиях. 15-дневные проростки переносили в полиэтиленовый пакет (Ш × Д × В; 4 × 4 × 12 см), содержащий садовую почву (как указано выше), и выращивали в контролируемых условиях окружающей среды (относительная влажность 80 ± 5%, 500–1000 мкмоль м –2 с –1 плотность потока фотосинтетических фотонов, 10 ± 2 hd –1 фотопериод от солнечного света и температуры окружающей среды 28 ± 2 ° C) в течение 7 недель.Эксперимент проводился в оранжерее Тайского научного парка, Патхум Тани, Таиланд (14 ° 01′12 ″ северной широты и 100 ° 31′12 ″ восточной долготы).
В качестве исходного материала были отобраны проростки однородного размера (высота растений 25 ± 2 см). Концентрация хлорида натрия (NaCl) в почвенном растворе была доведена до 0 (контроль), 200 (легкий стресс), 400 и 600 мМ (высокий стресс). Через 9 дней, признаки роста, содержание Na + , K + и Ca 2+ , содержание растворимого сахара, содержание свободного пролина, фотосинтетические пигменты, флуоресценция хлорофилла, чистая скорость фотосинтеза (P n ) были измерены устьичная проводимость (g с ) и скорость транспирации (E) в филлодиях ( n = 6).
Рост производительности
Измеряли высоту побегов (SH), длину корней (RL), количество филлод (NL), количество корней (NR), сырую массу (FW), сухую массу (DW) и площадь листовых филлод (LA) проростков. . Проростки акации сушили при 80 ° C в сушильном шкафу с горячим воздухом в течение 2 дней, а затем инкубировали в эксикаторе перед измерением DW. Площадь филлоды измеряли с использованием DT-сканирования измерителя площади корня / листа (Delta-Scan Version 2.03, Delta-T Devices, Ltd, Кембридж, Великобритания).
Na + , K + и Ca 2+ Анализ
Na + , K + и Ca 2+ анализировали в соответствии с модифицированным методом Tanaka et al.(1999) и Hossain et al. (2006). Вкратце, филлодные ткани проростков, подвергшихся солевому стрессу, промывали деионизированной водой для удаления поверхностного загрязнения Na + . Ткани измельчали в порошок в жидком азоте, экстрагировали кипящей дистиллированной водой и центрифугировали при 10000 × g в течение 10 мин. Супернатант фильтровали через мембранный фильтр 0,45 мкм (VertiPure ™ , Vertical ® ). Клеточные концентрации Na + , K + и Ca 2+ определяли с помощью ВЭЖХ (Waters Associates, Милфорд, Массачусетс, США) в сочетании с детектором проводимости 432 и ионно-эксклюзионной колонкой WATER IC-PACK ™ ( Waters Associates, Милфорд, Массачусетс, США).В качестве подвижной фазы использовали наночистую воду с минимальным расходом 0,6 мл –1 . В качестве стандарта использовали Na + , K + и Ca 2+ (Sigma, США).
Супернатант фильтровали через мембранный фильтр 0,45 мкм (VertiPure ™ , Vertical ® ). Клеточные концентрации Na + , K + и Ca 2+ определяли с помощью ВЭЖХ (Waters Associates, Милфорд, Массачусетс, США) в сочетании с детектором проводимости 432 и ионно-эксклюзионной колонкой WATER IC-PACK ™ ( Waters Associates, Милфорд, Массачусетс, США).В качестве подвижной фазы использовали наночистую воду с минимальным расходом 0,6 мл –1 . В качестве стандарта использовали Na + , K + и Ca 2+ (Sigma, США).
Определение общего количества растворимых сахаров
Общие растворимые сахара (сахароза, глюкоза и фруктоза) во втором полностью расширенном филлоде (от кончика побега) анализировали в соответствии с модифицированным методом Karkacier et al. (2003). В предварительно охлажденной ступке 100 мг ткани измельчали жидким азотом, экстрагировали 1 мл деионизированной воды, энергично встряхивали в течение 15 с, обрабатывали ультразвуком в течение 15 минут и затем центрифугировали при 12000 об / мин в течение 15 минут. Супернатант фильтровали через мембранный фильтр 0,45 мкм (VertiPure ™ , Vertical ® ) и хранили при –20 ° C до измерения общего содержания растворимых сахаров с помощью ВЭЖХ. Объем 40 мкл сырых экстрактов автоматически вводили в ВЭЖХ Waters, снабженную насосом Waters 600, с использованием колонки MetaCarb 87 ° C, снабженной защитной колонкой. В качестве подвижной фазы использовали деионизированную воду при расходе 0,5 мл мин. –1 . Обнаружение в режиме онлайн выполнялось с использованием детектора дифференциального рефрактометра Waters 410, а данные анализировались с помощью программного обеспечения Empower ® .В качестве стандартов использовали сахарозу, глюкозу и фруктозу (Fluka, США).
Супернатант фильтровали через мембранный фильтр 0,45 мкм (VertiPure ™ , Vertical ® ) и хранили при –20 ° C до измерения общего содержания растворимых сахаров с помощью ВЭЖХ. Объем 40 мкл сырых экстрактов автоматически вводили в ВЭЖХ Waters, снабженную насосом Waters 600, с использованием колонки MetaCarb 87 ° C, снабженной защитной колонкой. В качестве подвижной фазы использовали деионизированную воду при расходе 0,5 мл мин. –1 . Обнаружение в режиме онлайн выполнялось с использованием детектора дифференциального рефрактометра Waters 410, а данные анализировались с помощью программного обеспечения Empower ® .В качестве стандартов использовали сахарозу, глюкозу и фруктозу (Fluka, США).
Анализ свободного пролина
Свободный пролин во втором полностью развернутом филлоде (из кончика побега) экстрагировали и анализировали согласно методу Bates et al. (1973). Пятьдесят миллиграмм свежего материала растерли с жидким азотом в ступке. Порошок гомогената смешивали с 1 мл водной сульфосалициловой кислоты (3%, мас. / Об. ), и раствор фильтровали через фильтровальную бумагу (Whatman № 1, Англия).Экстрагированный раствор подвергали взаимодействию с равным объемом ледяной уксусной кислоты и реагента нингидрина (1,25 мг нингидрина в 30 мл ледяной уксусной кислоты и 20 мл 6 M H 3 PO 4 ) и инкубировали при 95 ° C в течение 1 часа. Реакцию останавливали, помещая контейнер в ледяную баню. Реакционную смесь энергично перемешивали с 2 мл толуола. После охлаждения до 25 ° C хромофор измеряли при 520 нм с помощью спектрофотометра UV-VIS (HACH DR / 4000; модель 48000, HACH Company, Loveland, CO, USA), используя L-пролин в качестве калибровочного стандарта.
Порошок гомогената смешивали с 1 мл водной сульфосалициловой кислоты (3%, мас. / Об. ), и раствор фильтровали через фильтровальную бумагу (Whatman № 1, Англия).Экстрагированный раствор подвергали взаимодействию с равным объемом ледяной уксусной кислоты и реагента нингидрина (1,25 мг нингидрина в 30 мл ледяной уксусной кислоты и 20 мл 6 M H 3 PO 4 ) и инкубировали при 95 ° C в течение 1 часа. Реакцию останавливали, помещая контейнер в ледяную баню. Реакционную смесь энергично перемешивали с 2 мл толуола. После охлаждения до 25 ° C хромофор измеряли при 520 нм с помощью спектрофотометра UV-VIS (HACH DR / 4000; модель 48000, HACH Company, Loveland, CO, USA), используя L-пролин в качестве калибровочного стандарта.
Фотосинтетические пигменты
Содержание хлорофилла а (Chl a ), хлорофилла b (Chl b ) и содержание TC во втором полностью развернутом филлоде анализировали по методу Shabala et al. (1998). 100 миллиграмм филлодной ткани гомогенизировали в стеклянных флаконах с использованием 10 мл 95,5% ацетона и смешивали с использованием гомогенизатора. Стеклянные флаконы герметизировали парафильмом ® для предотвращения испарения, а затем хранили при 4 ° C в течение 48 часов.Концентрации Chl a и Chl b измеряли при 662 нм и 644 нм с использованием спектрофотометра UV-VIS против ацетона (95,5%) в качестве холостого опыта.
100 миллиграмм филлодной ткани гомогенизировали в стеклянных флаконах с использованием 10 мл 95,5% ацетона и смешивали с использованием гомогенизатора. Стеклянные флаконы герметизировали парафильмом ® для предотвращения испарения, а затем хранили при 4 ° C в течение 48 часов.Концентрации Chl a и Chl b измеряли при 662 нм и 644 нм с использованием спектрофотометра UV-VIS против ацетона (95,5%) в качестве холостого опыта.
Флуоресценция хлорофилла
Излучение флуоресценции хлорофилла измеряли с адаксиальной поверхности второго полностью расширенного филлода с помощью системы мониторинга флуоресценции (FMS 2; Hansatech Instruments Ltd., Норфолк, Великобритания) в режиме модуляции амплитуды импульсов (Loggini et al., 1999).Филлод, выдержанный в темноте в течение 30 мин, первоначально подвергался воздействию модулированного измерительного луча дальнего красного света (светодиодный источник с типичным пиком на длине волны 735 нм). Исходный (F 0 ) и максимальный (F m ) выходы флуоресценции были измерены в слабомодулированном красном свете (<0,5 мкмоль м –2 с –1 ) с импульсами насыщающего света 1,6 с (> 6,8 мкмоль м –2 с –1 PAR) и рассчитано с использованием программного обеспечения FMS для Windows ® .Выход переменной флуоресценции (F v ) рассчитывали с использованием уравнения: F v = F m –F 0 . Отношение переменной флуоресценции к максимальной (F v / F m ) рассчитывали как максимальный квантовый выход фотохимии PSII. Выход фотонов ФСII (Φ PSII ) на свету был рассчитан как: Φ PSII = (F м ′ -F) / F m ′ через 45 с освещение, когда было достигнуто устойчивое состояние (Максвелл и Джонсон, 2000).
Исходный (F 0 ) и максимальный (F m ) выходы флуоресценции были измерены в слабомодулированном красном свете (<0,5 мкмоль м –2 с –1 ) с импульсами насыщающего света 1,6 с (> 6,8 мкмоль м –2 с –1 PAR) и рассчитано с использованием программного обеспечения FMS для Windows ® .Выход переменной флуоресценции (F v ) рассчитывали с использованием уравнения: F v = F m –F 0 . Отношение переменной флуоресценции к максимальной (F v / F m ) рассчитывали как максимальный квантовый выход фотохимии PSII. Выход фотонов ФСII (Φ PSII ) на свету был рассчитан как: Φ PSII = (F м ′ -F) / F m ′ через 45 с освещение, когда было достигнуто устойчивое состояние (Максвелл и Джонсон, 2000).
Чистая скорость фотосинтеза (P n ), устьичная проводимость (g s ) и скорость транспирации (E)
Чистая скорость фотосинтеза (P n ; мкмоль м –2 с –1 ), устьичная проводимость (г с ; мкмоль CO 2 м –2 с –1 ) и скорость транспирации (E; ммоль м –2 с –1 ) во втором полностью развернутом филлоде измеряли с помощью портативной системы фотосинтеза, оснащенной инфракрасным газоанализатором (модель LI 6400, LI-COR ® Inc. , Линкольн, штат Нью-Йорк, США). Согласно Cha-um et al., G s и E измерялись непрерывно путем мониторинга содержания воздуха, входящего и существующего в камере свободного пространства IRGA. (2007). Камера для образца была настроена на расход воздуха 500 мкмоль с –1 и температуру в камере 25 ° C. Интенсивность света была отрегулирована до 1000 мкмоль м –2 с –1 PPFD для красно-синего светодиодного источника света 6400-02B.
, Линкольн, штат Нью-Йорк, США). Согласно Cha-um et al., G s и E измерялись непрерывно путем мониторинга содержания воздуха, входящего и существующего в камере свободного пространства IRGA. (2007). Камера для образца была настроена на расход воздуха 500 мкмоль с –1 и температуру в камере 25 ° C. Интенсивность света была отрегулирована до 1000 мкмоль м –2 с –1 PPFD для красно-синего светодиодного источника света 6400-02B.
План эксперимента и статистический анализ
Эксперимент был организован как полностью рандомизированный блочный дизайн (CRBD) с шестью повторами ( n = 6).Полученные средние значения сравнивали с использованием HSD Тьюки и анализировали с помощью программного обеспечения SPSS.
Результаты
Морфологические показатели и показатели роста
Цвет листьев контрольных (0 мМ NaCl) и растений, подвергшихся слабому солевому стрессу (200 мМ NaCl), был зеленым. Напротив, филлоды и двупарчатые листья (настоящие старые листья) пожелтели у растений, выращенных в условиях экстремального солевого стресса (400–600 мМ NaCl) в течение 9 дней. Общие признаки роста снизились у растений, подвергшихся сильному солевому стрессу в течение 9 дней (рис. 1).SH, RL, NL, NR, FW, DW и филлодный LA у соленой акации, выращенной при стрессе 200 мМ NaCl, увеличились; однако они значительно уменьшились, когда растения подвергались воздействию 400–600 мМ NaCl в течение 9 дней (Таблица 1 и Рисунки 2A – C). SH, RL, NL, NR, FW побега и FW корня у растений с сильным солевым стрессом (600 мМ NaCl) резко снизились на 25, 23, 60, 45, 78 и 74%, соответственно, по сравнению с растениями, растущими под легкий стресс (200 мМ NaCl; Таблица 1). Кроме того, параметры чувствительности к соли, такие как NL, FW и DW, были снижены у растений, подвергшихся воздействию 400 мМ NaCl в течение 9 дней, аналогичный результат для 600 мМ.Рост и развитие соленой акации в условиях умеренного солевого стресса были усилены и лучше, чем у растений в контрольных условиях (0 мМ NaCl; Таблица 1).
Общие признаки роста снизились у растений, подвергшихся сильному солевому стрессу в течение 9 дней (рис. 1).SH, RL, NL, NR, FW, DW и филлодный LA у соленой акации, выращенной при стрессе 200 мМ NaCl, увеличились; однако они значительно уменьшились, когда растения подвергались воздействию 400–600 мМ NaCl в течение 9 дней (Таблица 1 и Рисунки 2A – C). SH, RL, NL, NR, FW побега и FW корня у растений с сильным солевым стрессом (600 мМ NaCl) резко снизились на 25, 23, 60, 45, 78 и 74%, соответственно, по сравнению с растениями, растущими под легкий стресс (200 мМ NaCl; Таблица 1). Кроме того, параметры чувствительности к соли, такие как NL, FW и DW, были снижены у растений, подвергшихся воздействию 400 мМ NaCl в течение 9 дней, аналогичный результат для 600 мМ.Рост и развитие соленой акации в условиях умеренного солевого стресса были усилены и лучше, чем у растений в контрольных условиях (0 мМ NaCl; Таблица 1).
Рисунок 1. Морфологические признаки растений проростков Acacia ampiceps , выращенных в условиях солевого стресса в течение 9 дней .
Таблица 1. Высота побега (SH), длина корня (RL), количество филлод (NL), количество корней (NR), сырой вес побегов (SHFW) и сырой вес корней (RTFW) Acacia ampiceps сеянцев, выращенных в условиях солевого стресса за 9 суток .
Рис. 2. Свежий вес (A), сухой вес (B), площадь листа филлода (C) и чистая скорость фотосинтеза (D) Acacia ampiceps проростков, выращенных в условиях солевого стресса в течение 9 дней. Данные представлены как среднее ± стандартная ошибка. Различные буквы означают значительную разницу при p ≤ 0,01 согласно тесту Тьюки HSD.
Na + , K + и Ca 2+
Содержание Na + и Ca 2+ в тканях филлод растений, подвергшихся солевому стрессу, постоянно увеличивалось, тогда как содержание K + значительно снижалось в зависимости от концентрации NaCl в почвенном растворе (Рисунки 3A – C). Na + / K + в листьях филлоды демонстрировал тенденцию, аналогичную обогащению Na, когда растения подвергались солевому стрессу (рис. 3D). В условиях экстремального солевого стресса (600 мМ NaCl) содержание Na + в тканях филлода достигло ∼33 мг г –1 DW, что привело к увеличению отношения Na + / K + на 7,8 раза (рисунок 3). Кроме того, содержание Na + и Ca 2+ в филлодных тканях растений с умеренным солевым стрессом (200 мМ NaCl) составляло 16.1 и 9,5 мг g –1 DW соответственно.
Na + / K + в листьях филлоды демонстрировал тенденцию, аналогичную обогащению Na, когда растения подвергались солевому стрессу (рис. 3D). В условиях экстремального солевого стресса (600 мМ NaCl) содержание Na + в тканях филлода достигло ∼33 мг г –1 DW, что привело к увеличению отношения Na + / K + на 7,8 раза (рисунок 3). Кроме того, содержание Na + и Ca 2+ в филлодных тканях растений с умеренным солевым стрессом (200 мМ NaCl) составляло 16.1 и 9,5 мг g –1 DW соответственно.
Рисунок 3. Содержание Na + (A), K + (B) и Ca 2+ (C) и соотношение Na + / K + (D) в амплицепсах Acacia сеянцев, выращенных в условиях солевого стресса в течение 9 дней. Данные представлены как среднее ± стандартная ошибка. Различные буквы означают значительную разницу при p ≤ 0,01 согласно тесту Тьюки HSD.
Свободный пролин и общий растворимый сахар
Содержание свободного пролина в филодах растений, подвергшихся солевому стрессу, значительно увеличилось, особенно у растений, находящихся в условиях экстремального стресса.Оно увеличивалось на 7,1 (13,18 мкмоль г –1 FW) и 28,7 раза (53,33 мкмоль г –1 FW) у растений, растущих под 400 и 600 мМ NaCl, по сравнению с контрольными растениями (1,86 мкмоль г -1 FW; Рисунок 4A). Точно так же общий растворимый сахар непрерывно накапливался в филодах растений, подвергшихся солевому стрессу, и достигал 42,1 мг / г –1 DW (~ 3,2 раза от контроля) у растений, выращенных под 600 мМ NaCl (рис. 4B). Содержание сахарозы, глюкозы и фруктозы у растений, подвергшихся солевому стрессу, увеличивалось в зависимости от степени солевого стресса.Сахароза, глюкоза и фруктоза в растениях, выращенных в среде 600 мМ NaCl, были в 2,2, 3,7 и 5,1 раза выше контроля, соответственно (таблица 2). Накопление глюкозы и фруктозы было сравнительно меньше у растений с концентрацией NaCl 200 мМ, где оно увеличивалось в 2,3 и 3,6 раза соответственно по сравнению с контрольными растениями.
Рисунок 4. Содержание свободного пролина (A) и общего растворимого сахара (B) в проростках Acacia ampiceps , выращенных в условиях солевого стресса в течение 9 дней. Данные представлены как среднее ± стандартная ошибка. Различные буквы означают значительную разницу при p ≤ 0,01 согласно тесту Тьюки HSD.
Таблица 2. Сахароза (Suc), глюкоза (Gluc), фруктоза (Fruc), устьичная проводимость (gs) и скорость транспирации (E) Acacia ampiceps проростки, выращенные в условиях солевого стресса в течение 9 дней .
Фотосинтетические способности
Chl a , Chl b , и TC в филлодиях были увеличены у растений, выращенных в условиях умеренного солевого стресса (200 мМ NaCl; таблица 3).Однако Chl a , Chl b и TC разлагались на 48,3, 29,9 и 39,8% контроля, соответственно, при экстремальном солевом стрессе (600 мМ NaCl; Таблица 3). Кроме того, Chl a , чувствительный параметр, значительно снижался, когда растения подвергались воздействию 400 мМ NaCl, тогда как Chl b сохранялся. Максимальный квантовый выход ФСII (F v / F m ) и выход фотонов ФСII (Ф ФСII ) снижались, когда растения подвергались сильному солевому стрессу (400–600 мМ NaCl) на 9 дней.При экстремальном солевом стрессе F v / F m и Φ PSII были уменьшены на 17,0 и 13,5%, соответственно, по сравнению с таковыми у растений с умеренным солевым стрессом (Таблица 3). Чистая скорость фотосинтеза (P n ) у растений с умеренным солевым стрессом была увеличена, а затем значительно снизилась при более сильном солевом стрессе в зависимости от концентрации (Рисунок 2D). Точно так же устьичная проводимость (g s ) и скорость транспирации (E) у растений с умеренным солевым стрессом достигли пика до 10.
Кроме того, Chl a , чувствительный параметр, значительно снижался, когда растения подвергались воздействию 400 мМ NaCl, тогда как Chl b сохранялся. Максимальный квантовый выход ФСII (F v / F m ) и выход фотонов ФСII (Ф ФСII ) снижались, когда растения подвергались сильному солевому стрессу (400–600 мМ NaCl) на 9 дней.При экстремальном солевом стрессе F v / F m и Φ PSII были уменьшены на 17,0 и 13,5%, соответственно, по сравнению с таковыми у растений с умеренным солевым стрессом (Таблица 3). Чистая скорость фотосинтеза (P n ) у растений с умеренным солевым стрессом была увеличена, а затем значительно снизилась при более сильном солевом стрессе в зависимости от концентрации (Рисунок 2D). Точно так же устьичная проводимость (g s ) и скорость транспирации (E) у растений с умеренным солевым стрессом достигли пика до 10. 44 мкмоль H 2 O m -2 s -1 и 3,13 ммоль H 2 O m -2 s -1 , тогда как они снизились на 73,3 и 73,5% соответственно, когда растения были подвергали воздействию 600 мМ NaCl (таблица 2).
44 мкмоль H 2 O m -2 s -1 и 3,13 ммоль H 2 O m -2 s -1 , тогда как они снизились на 73,3 и 73,5% соответственно, когда растения были подвергали воздействию 600 мМ NaCl (таблица 2).
Таблица 3. Хлорофилл а (Chl a ), хлорофилл b (Chl b ) и общая концентрация хлорофилла (TC), максимальный квантовый выход ФСII (F v / F m ) и выход фотонов ФСII (Φ ФСII ) проростков Acacia ampiceps , выращенных в условиях солевого стресса в течение 9 дней .
Связь между биохимическими и физиологическими изменениями
Обогащение Na + в филлодных тканях растений, подвергшихся солевому стрессу, было положительно связано с деградацией TC ( R 2 = 0,72; фиг. 5A). Следовательно, деградация Chl a положительно коррелировала со снижением максимального квантового выхода ФСII (F v / F m ) ( R 2 = 0,59; Рисунок 5B).Точно так же уменьшение выхода фотонов ФСII (Φ PSII ) было сильно связано со снижением чистой скорости фотосинтеза (P n ) ( R 2 = 0,81; Рисунок 5C), что привело к уменьшению растений. DW ( R 2 = 0,91; рисунок 5D).
5A). Следовательно, деградация Chl a положительно коррелировала со снижением максимального квантового выхода ФСII (F v / F m ) ( R 2 = 0,59; Рисунок 5B).Точно так же уменьшение выхода фотонов ФСII (Φ PSII ) было сильно связано со снижением чистой скорости фотосинтеза (P n ) ( R 2 = 0,81; Рисунок 5C), что привело к уменьшению растений. DW ( R 2 = 0,91; рисунок 5D).
Рис. 5. Зависимость между (A) Na + и общим содержанием хлорофилла, (B) содержанием хлорофилла a и максимальным квантовым выходом ФСII (F v / F m ) , (C) выход фотонов PSII (Φ PSII ) и чистая скорость фотосинтеза (P n ), и (D) чистая скорость фотосинтеза и сухой вес растения Acacia ampiceps саженцев выращены в условиях солевого стресса за 9 суток. Планки погрешностей представляют ± SE.
Планки погрешностей представляют ± SE.
Обсуждение
Показатели роста A. ampiceps лучше проявлялись при низкой концентрации соли; однако он снижался в зависимости от степени солевого загрязнения почвы. Ранее сообщалось, что процент выживания A. ampiceps был максимальным (98,1%) с SH 127,2 см при выращивании в среде обитания с засоленной почвой в течение 9 месяцев (Shirazi et al., 2006). Смертность A. ampiceps , выращенных в гидропонной культуре, составляла всего 7% (сохраняя 38% DW от контроля), когда растения подвергались воздействию 1.5% NaCl (257 мМ) и увеличился до 47% (только с 22% DW контроля) при 2,0% NaCl (342 мМ) (Yokota, 2003). Наши результаты подтверждаются более ранним исследованием, в котором сообщалось об улучшении характеристик роста, таких как FW побега, DW для корня, FW и DW- корня A. ampiceps при умеренном солевом стрессе (EC e = 10 dS m — 1 ) в гидропонной культуре, а затем снизился при сильном солевом стрессе (EC e = 20–30 dS m –1 ), особенно при pH 9. 5 (Ажар и др., 2013). Напротив, SH, RL, FW побега, DW побега, FW корня и DW корня A. ampiceps снижались у растений, подвергшихся воздействию 100–300 мМ NaCl в течение 2 дней (Abbas et al., 2013). Кроме того, Marcar и Crawford (2011) наблюдали снижение SH в нескольких источниках происхождения A. ampiceps при 150 моль / м –3 солевой обработки и в условиях солевого заболачивания. Параллельно с нашими выводами, SH и диаметр ствола A. ampiceps были увеличены в полевых испытаниях в Эр-Рияде, что привело к увеличению конечного выхода биомассы на 39.69 тонн га –1 (Aref et al., 2003). На засоленных полях (EC e = 13,9 dS m –1 ) SH и количество ветвей на растение были увеличены в зависимости от периода культивирования (6–24 месяца; Abbas et al., 2015). Количество листьев — хороший показатель для определения смертности растений, выращенных в условиях солевого стресса. Махмуд и др. (2009) предположили, что количество листьев A.
5 (Ажар и др., 2013). Напротив, SH, RL, FW побега, DW побега, FW корня и DW корня A. ampiceps снижались у растений, подвергшихся воздействию 100–300 мМ NaCl в течение 2 дней (Abbas et al., 2013). Кроме того, Marcar и Crawford (2011) наблюдали снижение SH в нескольких источниках происхождения A. ampiceps при 150 моль / м –3 солевой обработки и в условиях солевого заболачивания. Параллельно с нашими выводами, SH и диаметр ствола A. ampiceps были увеличены в полевых испытаниях в Эр-Рияде, что привело к увеличению конечного выхода биомассы на 39.69 тонн га –1 (Aref et al., 2003). На засоленных полях (EC e = 13,9 dS m –1 ) SH и количество ветвей на растение были увеличены в зависимости от периода культивирования (6–24 месяца; Abbas et al., 2015). Количество листьев — хороший показатель для определения смертности растений, выращенных в условиях солевого стресса. Махмуд и др. (2009) предположили, что количество листьев A. ampiceps было уменьшено на 10, 21 и 37% по сравнению с контролем при воздействии EC e 10, 20 и 30 dS m –1 соответственно на 2 года.
ampiceps было уменьшено на 10, 21 и 37% по сравнению с контролем при воздействии EC e 10, 20 и 30 dS m –1 соответственно на 2 года.
В нашем исследовании Na + и Ca 2+ в филлодах A. ampiceps постоянно обогащались в зависимости от концентрации соли в почвенном растворе, тогда как K + снижалось, что приводило к увеличению Na . + / K + соотношение. У галофитов Na + был обогащен органеллами вакуолей, что связано с активностью Na + / H + антипортера (NHXs) и локализации вакуолярной H + -АТФазы (VH + -ATPase) на вакуолярной мембрана (Barkla et al., 2002; Wang et al., 2015). Это соответствует предыдущим выводам Marcar et al. (1991) и Аббас и др. (2013), которые сообщили об увеличении соотношения Na: K или снижении соотношения K: Na соответственно. Содержание Na + в новом филлоде (молодом) A. ampiceps , выращенном в условиях солевого стресса (EC e = 40 dS m –1 в летний сезон) было максимальным и больше, чем в более старых филлодах ( Ширази и др. , 2001). Аналогичное наблюдение (накопление Na + и восстановление K + ) было продемонстрировано на других видах акации ( A.auriculiformis , A. mangium , A. longifolia и A. senegal ), выращенных в условиях солевого стресса (Marcar et al., 1991; Hardikar, Pandey, 2008; Morais et al., 2012). Sunarpi et al. (2005) предположили, что высокие концентрации соли вызывают поглощение и транспорт Na + и снижают содержание K + в побегах. В вакуолях K + играют ключевую роль в генерации тургора; однако во время засоления происходит выброс K + и приток Na + (Pottosin and Maiz, 2002; Adams and Shin, 2014).Несмотря на это, было обнаружено, что Na + помогает в поддержании тургора (Kronzucker et al., 2013).
, 2001). Аналогичное наблюдение (накопление Na + и восстановление K + ) было продемонстрировано на других видах акации ( A.auriculiformis , A. mangium , A. longifolia и A. senegal ), выращенных в условиях солевого стресса (Marcar et al., 1991; Hardikar, Pandey, 2008; Morais et al., 2012). Sunarpi et al. (2005) предположили, что высокие концентрации соли вызывают поглощение и транспорт Na + и снижают содержание K + в побегах. В вакуолях K + играют ключевую роль в генерации тургора; однако во время засоления происходит выброс K + и приток Na + (Pottosin and Maiz, 2002; Adams and Shin, 2014).Несмотря на это, было обнаружено, что Na + помогает в поддержании тургора (Kronzucker et al., 2013).
Более ранние исследования продемонстрировали, что галофиты обладают конститутивными антипортерами тонопластов, которые способствуют накоплению Na + в вакуоли через систему регуляции SOS (Blumwald, 2000). В молекулярно-биологических исследованиях NHXs, вакуолярные антипортеры Na + / H + , чтобы загнать Na + в вакуоль галофитных видов, Aeluropus littoralis (Zhang et al., 2008) и Thellungiella halophila (Vera-Estrella et al., 2005). NHX, происходящие от видов галофитов, были охарактеризованы, клонированы и трансформированы в табак (Jha et al., 2011), Arabidopsis (Li et al., 2007) и рисовые культуры (Verma et al., 2007). Мы обнаружили повышение Ca 2+ в филлоде A. ampiceps , выращенных в условиях солевого стресса, и это резко контрастировало с пониженным содержанием Ca 2+ у других видов Acacia , когда растения подвергались солевому стрессу ( Rehman et al., 2000; Хардикар и Пандей, 2008; Patel et al., 2010). Ширази и др. (2001) сообщили, что накопление Ca 2+ достигло пика при EC e = 40 dS m –1 в летний сезон, и оно было в 1,47 раза больше, чем у растений с низким EC e .
В молекулярно-биологических исследованиях NHXs, вакуолярные антипортеры Na + / H + , чтобы загнать Na + в вакуоль галофитных видов, Aeluropus littoralis (Zhang et al., 2008) и Thellungiella halophila (Vera-Estrella et al., 2005). NHX, происходящие от видов галофитов, были охарактеризованы, клонированы и трансформированы в табак (Jha et al., 2011), Arabidopsis (Li et al., 2007) и рисовые культуры (Verma et al., 2007). Мы обнаружили повышение Ca 2+ в филлоде A. ampiceps , выращенных в условиях солевого стресса, и это резко контрастировало с пониженным содержанием Ca 2+ у других видов Acacia , когда растения подвергались солевому стрессу ( Rehman et al., 2000; Хардикар и Пандей, 2008; Patel et al., 2010). Ширази и др. (2001) сообщили, что накопление Ca 2+ достигло пика при EC e = 40 dS m –1 в летний сезон, и оно было в 1,47 раза больше, чем у растений с низким EC e . Ca 2+ снижает засоление, защищает растения и придает устойчивость к засолению гликофитов (Greenway and Munns, 1980; Hadi and Karimi, 2012). Сообщалось, что экзогенный Ca 2+ снижает засоление галофитов (Grigore et al., 2012). Ca 2+ играет ключевую роль в стабилизации структуры клеточной стенки, поддержании структурной и функциональной целостности клеточной мембраны, регуляции ионного транспорта и ионообменном поведении, помимо того, что он действует как посредник в передаче сигналов стресса (Hasegawa et al., 2000; Хади, Карими, 2012). Мы наблюдали обогащение Ca 2+ и Na + в условиях экстремальной солености. Повышенное накопление Ca 2+ , несмотря на обогащение Na + , резко контрастировало с тем, что Na + препятствует поглощению Ca 2+ при солевом стрессе (Rehman et al., 2000). Накопление Ca 2+ в экстремальных солевых условиях может защитить A. ampiceps от токсического действия Na + путем активации путей SOS, которые защищают клеточные мембраны от повреждения, вызванного засолением.
Ca 2+ снижает засоление, защищает растения и придает устойчивость к засолению гликофитов (Greenway and Munns, 1980; Hadi and Karimi, 2012). Сообщалось, что экзогенный Ca 2+ снижает засоление галофитов (Grigore et al., 2012). Ca 2+ играет ключевую роль в стабилизации структуры клеточной стенки, поддержании структурной и функциональной целостности клеточной мембраны, регуляции ионного транспорта и ионообменном поведении, помимо того, что он действует как посредник в передаче сигналов стресса (Hasegawa et al., 2000; Хади, Карими, 2012). Мы наблюдали обогащение Ca 2+ и Na + в условиях экстремальной солености. Повышенное накопление Ca 2+ , несмотря на обогащение Na + , резко контрастировало с тем, что Na + препятствует поглощению Ca 2+ при солевом стрессе (Rehman et al., 2000). Накопление Ca 2+ в экстремальных солевых условиях может защитить A. ampiceps от токсического действия Na + путем активации путей SOS, которые защищают клеточные мембраны от повреждения, вызванного засолением. Но как это происходит в A. ampiceps , пока неизвестно.
Но как это происходит в A. ampiceps , пока неизвестно.
Свободный пролин и общий растворимый сахар в филодах A. ampiceps , выращенных в условиях солевого стресса, были обогащены пропорционально концентрации соли в почве. Это предполагает положительную роль пролина в регулировании осмотического давления при солевом стрессе.Эти данные подтверждаются аналогичным наблюдением, сделанным ранее на тканях листьев и корней A. ampiceps , A . mangium и A. auriculiformis , выращенные в условиях солевого стресса в теплице, в зависимости от степени концентрации NaCl или EC e (Diouf et al., 2005; Patel et al., 2010). Галофиты накапливают больше пролина, чем гликофиты, и это связано с подавлением катаболизма пролина пролин-окисляющим ферментом, PDH (пролиндегидрогеназой), и усиленным синтезом пролина через пирролин-5-карбоксилатсинтетазу (P5CS; Taji et al., 2004; Кант и др., 2006).
Ранее исследования показали, что общее содержание растворимого сахара, то есть сахарозы, глюкозы и фруктозы, было повышено в пальме Nipa, подвергшейся воздействию 16,6 dS m –1 EC e в качестве стресса NaCl (Theerawitaya et al. , 2014) и в семядолях квиноа галофитов, подвергнутых воздействию 200 мМ NaCl в течение 4 дней (Rosa et al., 2009a). Сахароза, основной источник углерода в растущих проростках, наряду с глюкозой действует как осмопротектор и помогает поддерживать гомеостаз и целостность биомембран во время абиотических стрессов (Rosa et al., 2009b). В нашем исследовании содержание сахара положительно коррелировало с уровнями Na + в филлодиях, тем самым предполагая роль в осмотическом регулировании на клеточном уровне в механизме солевой защиты в качестве осмопротектора (Ghosh et al., 2001). В растениях сахара не только функционируют как субстраты для энергетического и углеродного метаболизма, но также действуют как осмолиты и посредники в передаче сигналов и модулируют рост, развитие и экспрессию генов (Rolland et al., 2002).
, 2014) и в семядолях квиноа галофитов, подвергнутых воздействию 200 мМ NaCl в течение 4 дней (Rosa et al., 2009a). Сахароза, основной источник углерода в растущих проростках, наряду с глюкозой действует как осмопротектор и помогает поддерживать гомеостаз и целостность биомембран во время абиотических стрессов (Rosa et al., 2009b). В нашем исследовании содержание сахара положительно коррелировало с уровнями Na + в филлодиях, тем самым предполагая роль в осмотическом регулировании на клеточном уровне в механизме солевой защиты в качестве осмопротектора (Ghosh et al., 2001). В растениях сахара не только функционируют как субстраты для энергетического и углеродного метаболизма, но также действуют как осмолиты и посредники в передаче сигналов и модулируют рост, развитие и экспрессию генов (Rolland et al., 2002).
Na + компартментация в органеллах вакуолей мангровых видов, Bruguiera sexangula было подтверждено накоплением ионов, связанным с увеличением размера вакуолей и регулированием активности Na + / H + обменника и H + -АТФаза (Kura-Hotta et al. , 2001; Мимура и др., 2003). В случае умеренного солевого стресса, компартментация Na + в A. ampiceps может играть ключевую роль в качестве механизма солевой защиты для уменьшения симптомов ионного отравления. Таким образом, ионная токсичность была обнаружена в более старом филлоде, то есть хлорозе A. ampiceps при воздействии 600 мМ NaCl в течение 9 дней. В мангровых зарослях Bruguiera parviflora ультраструктура хлоропластов в растении, выращенном под 400 мМ NaCl, была разрушена, а также снижены как содержание, так и активность пигментных белковых и тилакоидных белковых комплексов в зависимости от степени солевой обработки (Parida et al. ., 2003), что привело к снижению чистой скорости фотосинтеза (Li et al., 2008). В настоящем исследовании пигмент хлорофилла, флуоресценция хлорофилла и чистая скорость фотосинтеза A. ampiceps были увеличены при слабом солевом стрессе (200 мМ NaCl), а затем снизились при сильном солевом стрессе (≥400 NaCl).
, 2001; Мимура и др., 2003). В случае умеренного солевого стресса, компартментация Na + в A. ampiceps может играть ключевую роль в качестве механизма солевой защиты для уменьшения симптомов ионного отравления. Таким образом, ионная токсичность была обнаружена в более старом филлоде, то есть хлорозе A. ampiceps при воздействии 600 мМ NaCl в течение 9 дней. В мангровых зарослях Bruguiera parviflora ультраструктура хлоропластов в растении, выращенном под 400 мМ NaCl, была разрушена, а также снижены как содержание, так и активность пигментных белковых и тилакоидных белковых комплексов в зависимости от степени солевой обработки (Parida et al. ., 2003), что привело к снижению чистой скорости фотосинтеза (Li et al., 2008). В настоящем исследовании пигмент хлорофилла, флуоресценция хлорофилла и чистая скорость фотосинтеза A. ampiceps были увеличены при слабом солевом стрессе (200 мМ NaCl), а затем снизились при сильном солевом стрессе (≥400 NaCl). Пигменты Chl a и Chl b пигментов A. auriculiformis , подвергнутых солевому стрессу, разложились в зависимости от уровня солености (в диапазоне от 1.От 2 до 10 дСм м –1 ). Параллельно с нашими выводами, F v / F m , Φ PSII , P n , g s , и E в пальме Nipa (галофитный вид) были вызваны низкой концентрацией соли (EC e = 8,9 dS m -1 ), а затем значительно упали, особенно при высокой концентрации (EC e = 57,2 dS m -1 ) ( Theerawitaya et al., 2014).
Пигменты Chl a и Chl b пигментов A. auriculiformis , подвергнутых солевому стрессу, разложились в зависимости от уровня солености (в диапазоне от 1.От 2 до 10 дСм м –1 ). Параллельно с нашими выводами, F v / F m , Φ PSII , P n , g s , и E в пальме Nipa (галофитный вид) были вызваны низкой концентрацией соли (EC e = 8,9 dS m -1 ), а затем значительно упали, особенно при высокой концентрации (EC e = 57,2 dS m -1 ) ( Theerawitaya et al., 2014).
Заключение
Na + и Ca 2+ в филлодах A. ampiceps при экстремальном солевом стрессе (600 мМ NaCl) были обогащены, в то время как K + снизился, что привело к увеличению отношения Na: K. Накопление Na + в филлодных тканях растений, подвергшихся солевому стрессу, может накапливаться в вакуолярных органеллах как компартментация Na + . С другой стороны, свободный пролин и растворимый сахар были увеличены на клеточных уровнях тканей листа, чтобы играть ключевую роль в осморегуляции механизма защиты от соли для защиты A.ampiceps от токсичности, вызванной солью. Следовательно, очевидно наблюдались деградация хлорофилла, уменьшение флуоресценции хлорофилла и общее снижение скорости фотосинтеза у A. ampiceps при сильном солевом стрессе, что приводило к общему подавлению роста.
С другой стороны, свободный пролин и растворимый сахар были увеличены на клеточных уровнях тканей листа, чтобы играть ключевую роль в осморегуляции механизма защиты от соли для защиты A.ampiceps от токсичности, вызванной солью. Следовательно, очевидно наблюдались деградация хлорофилла, уменьшение флуоресценции хлорофилла и общее снижение скорости фотосинтеза у A. ampiceps при сильном солевом стрессе, что приводило к общему подавлению роста.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось в отсутствие каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Это исследование было поддержано Национальным центром генной инженерии и биотехнологии и частично профинансировано Национальным агентством развития науки и технологий (NSTDA) для постдокторской должности в CT и программы стипендий TWAS для HPS. Мы также благодарим Департамент землеустройства Министерства сельского хозяйства и кооперативов Таиланда за предоставление семян Acacia ampiceps для исследования.
Список литературы
Аббас, Г., Сакиб, М., Ахтар, Дж., И Басра, С. М. А. (2013). Потенциал устойчивости к засолению двух видов Acacia на ранней стадии всходов. Пак. J. Agric Sci. 50, 683–688.
Google Scholar
Аббас, Г., Сакиб, М., Ахтар, Дж., Муртаза, Г., Шахид, М., и Хуссейн, А. (2015). Связь между подкислением ризосферы и фиторемедиацией у двух видов Acacia . J. Soil Sedim. DOI: 10.1007 / s11368-014-1051-9
CrossRef Полный текст | Google Scholar
Адамс, Э., и Шин, Р. (2014). Транспорт, передача сигналов и гомеостаз калия и натрия в растениях. J. Integr. Plant Biol. 56, 231–249.
Google Scholar
Ансари Р., Хан М. А. и Гул Б. (2007). «Выгодное использование засоленных земель: перспективы и меры предосторожности», в Растениеводство и кормопроизводство с использованием соленой воды , Vol. 10, ред. М. Кафи и М. А. Хан (Нью-Дели: NAM S&T Center), 103–108.
Google Scholar
Ареф, И. М., Эль-Джухани, Л. И., и Хегази, С. С. (2003). Сравнение роста и производства биомассы шести видов Acacia в Эр-Рияде, Саудовская Аравия, после 4 лет орошаемого выращивания. J. Arid. Environ. 54, 783–792. DOI: 10.1006 / jare.2002.1067
М., Эль-Джухани, Л. И., и Хегази, С. С. (2003). Сравнение роста и производства биомассы шести видов Acacia в Эр-Рияде, Саудовская Аравия, после 4 лет орошаемого выращивания. J. Arid. Environ. 54, 783–792. DOI: 10.1006 / jare.2002.1067
CrossRef Полный текст | Google Scholar
Ашраф, М. А., Аван, А. Р., и Махмуд, К. (2012). Восстановление засоленных экосистем путем выращивания солеустойчивых растений. Пак. J. Bot. 44, 69–75.
Google Scholar
Азхар, М. Ф., Шах, А., Х., Сиддиуи, М. А., и Анджум, К. (2013). Показатели роста Acacia ampiceps Maslin при комбинированном воздействии различных уровней солености и pH. World Appli. Sci. J. 28, 562–567.
Google Scholar
Баркла, Б. Дж., Вера-Эстрелла, Р., Камачо-Эмитерио, Дж., И Пантоха, О. (2002). Обмен Na + / H + в галофите Mesembryanthemum crisinum связан с клеточными сайтами хранения Na + . Функц. Plant Biol. 29, 1017–1024. DOI: 10.1071 / FP02045
29, 1017–1024. DOI: 10.1071 / FP02045
CrossRef Полный текст | Google Scholar
Бейтс, Л. С., Уолдрен, Р. П., и Тир, И. Д. (1973). Быстрое определение свободного пролина для исследований водного стресса. Почва растений 39, 205–207. DOI: 10.1007 / BF00018060
CrossRef Полный текст | Google Scholar
Ча-ум, С., Супайбулватана, К., и Кирдмани, К. (2007). Накопление глицинбетаина, физиологические характеристики и эффективность роста солеустойчивых и солеочувствительных линий риса индика ( Oryza sativa L.ssp. индика) в ответ на солевой стресс. J. Agron. Crop Sci. 193, 157–166. DOI: 10.1111 / j.1439-037X.2007.00251.x
CrossRef Полный текст | Google Scholar
Диуф, Д., Дюпионуа, Р., Ба, А. Т., Нейра, М., и Лесюер, Д. (2005). Симбиоз Acacia auriculiformis и Acacia mangium с микоризными грибами и Bradyrhizobium spp. улучшает солеустойчивость в тепличных условиях. Функц. Plant Biol. 32, 1143–1152.DOI: 10.1071 / FP04069
32, 1143–1152.DOI: 10.1071 / FP04069
CrossRef Полный текст | Google Scholar
Дайнс, Р.А., и Шлинк, А.С. (2002). Потенциал животноводства Австралии вида Acacia . Консерв. Sci. 4, 117–124.
Google Scholar
Флауэрс, Т. Дж., Галал, Х. К. и Бромхэм Л. (2010). Эволюция галофитов: множественные причины солеустойчивости наземных растений. Функц. Plant Biol. 37, 604–612. DOI: 10.1071 / FP09269
CrossRef Полный текст | Google Scholar
Цветы, Т.Дж., Хаджибагери М. А. и Клипсон Н. Дж. У. (1986). Галофиты. Q. Rev. Biol. 61, 313–337. DOI: 10.1086 / 415032
CrossRef Полный текст | Google Scholar
Гош С., Багчи С. и Маджумдер А. Л. (2001). Фруктозо-1,6-бифосфатаза хлоропластов из Oryza отличается свойством солеустойчивости от фермента Porteresia и защищена осмолитами. Plant Sci. 160, 1171–1181. DOI: 10.1016 / S0168-9452 (01) 00361-2
CrossRef Полный текст | Google Scholar
Гири, Б. , Капур Р. и Мукерджи К. Г. (2003). Влияние грибов арбускулярной микориз и засоления на рост, биомассу и минеральное питание Acacia auriculiformis . Biol. Fertil. Почва 38, 170–175. DOI: 10.1007 / s00374-003-0636-z
, Капур Р. и Мукерджи К. Г. (2003). Влияние грибов арбускулярной микориз и засоления на рост, биомассу и минеральное питание Acacia auriculiformis . Biol. Fertil. Почва 38, 170–175. DOI: 10.1007 / s00374-003-0636-z
CrossRef Полный текст | Google Scholar
Гринуэй, М., и Маннс, Р. (1980). Механизмы солеустойчивости негалофитов. Ann. Rev. Plant Physiol. 31, 149–190. DOI: 10.1146 / annurev.pp.31.060180.001053
CrossRef Полный текст | Google Scholar
Григоре, М.Н., Боскайу М., Ллинарес Дж. И Висенте О. (2012). Смягчение индуцированного солевым стрессом подавления репродуктивного развития Plantago crassifolia с помощью дополнительных препаратов кальция или магния. Нет. Бот. Horti Agrobot. Клуж-Нап. 40, 58–66.
Google Scholar
Хади, М. Р., Карими, Н. (2012). Роль кальция в солеустойчивости растений. J. Plant Nutr. 35, 2037–2054. DOI: 10.1080 / 017.2012.717158
CrossRef Полный текст | Google Scholar
Хардикар, С. А., Панди А. Н. (2008). Рост, водный статус и накопление питательных веществ сеянцев Acacia senegal (L.) Willd. в ответ на засоление почвы. Анал. Биол. 30, 17–28.
А., Панди А. Н. (2008). Рост, водный статус и накопление питательных веществ сеянцев Acacia senegal (L.) Willd. в ответ на засоление почвы. Анал. Биол. 30, 17–28.
Google Scholar
Hasegawa, P.H., Bressan, R.A., Zhu, J.K., and Bohnert, H.J. (2000). Клеточные и молекулярные реакции растений на высокое засоление. Ann. Rev. Plant Physiol. Завод Мол. Биол. 51, 463–499. DOI: 10.1146 / annurev.arplant.51.1.463
CrossRef Полный текст | Google Scholar
Хоссейн, Г.С., Вадитее, Р., Хибино, Т., Танака, Ю., и Такабе, Т. (2006). Корневоспецифическая экспрессия гена-антипортера Na + / H + из Synechocystis sp. PCC6803 придает солеустойчивость табачного растения. Plant Biotechnol. 23, 275–281. DOI: 10.5511 / plantbiotechnology.23.275
CrossRef Полный текст | Google Scholar
Джа А., Джоши М., Ядав Н. С., Агарвал П. К. и Джа Б. (2011). Клонирование и характеристика антипортерного гена SbNHX1 Salicornia brachiata Na + / H + и его экспрессия при абиотическом стрессе. Мол. Биол. Rep. 38, 1965–1973. DOI: 10.1007 / s11033-010-0318-5
Мол. Биол. Rep. 38, 1965–1973. DOI: 10.1007 / s11033-010-0318-5
CrossRef Полный текст | Google Scholar
Кант, С., Кант, П., Раве, Э., и Барак, С. (2006). Доказательства того, что дифференциальная экспрессия гена между галофитами, Thellungiella halophila и Arabidopsis thaliana отвечает за более высокие уровни совместимого осмолита пролина и жесткий контроль захвата Na + в T. halophila . Plant Cell Environ. 29, 1220–1234. DOI: 10.1111 / j.1365-3040.2006.01502.x
CrossRef Полный текст | Google Scholar
Каркасьер М., Эбрас М., Услу М. К. и Аксу М. (2003). Сравнение различных методов экстракции и обнаружения сахаров с использованием ВЭЖХ с аминосвязанной фазой. J. Chromatogr. Sci. 41, 331–333. DOI: 10.1093 / chromsci / 41.6.331
CrossRef Полный текст | Google Scholar
Кронцукер, Х. Дж., Коскун, Д., Шульце, Л. М., Вонг, Дж. Р., и Бритто, Д.Т. (2013). Натрий как питательное и токсичное вещество. Почва растений 369, 1–23. DOI: 10.1007 / s11104-013-1801-2
Почва растений 369, 1–23. DOI: 10.1007 / s11104-013-1801-2
CrossRef Полный текст | Google Scholar
Кура-Хотта, М., Мимура, М., Цудзимура, Т., Вашитани-Немото, С., и Мимура, Т. (2001). Экструзия Na + , вызванная высокой солевой обработкой, и индуцированная низкой солевой обработкой накопление Na + в культивируемых в суспензии клетках мангрового растения, Bruguiera sexangula . Plant Cell Environ. 24, 1105–1112.DOI: 10.1046 / j.0016-8025.2001.00761.x
CrossRef Полный текст | Google Scholar
Ли, Дж., Цзян, Г., Хуанг, П., Ма, Дж. И Чжан, Ф. (2007). Сверхэкспрессия гена-антипортера Na + / H + из Suaeda salsa придает устойчивость к холоду и соли трансгенному Arabidopsis thaliana . Растительная клеточная ткань. Орг. Культ. 90, 41–48. DOI: 10.1007 / s11240-007-9246-z
CrossRef Полный текст | Google Scholar
Ли, Н., Chen, S., Zhou, X., Li, C. , Shao, J., Wang, R., et al. (2008). Влияние NaCl на фотосинтез, накопление соли и компартментацию у двух видов мангровых зарослей, Kandelia candel и Bruguiera gymnorrhiza . Aqua. Бот. 88, 303–310. DOI: 10.1016 / j.aquabot.2007.12.003
, Shao, J., Wang, R., et al. (2008). Влияние NaCl на фотосинтез, накопление соли и компартментацию у двух видов мангровых зарослей, Kandelia candel и Bruguiera gymnorrhiza . Aqua. Бот. 88, 303–310. DOI: 10.1016 / j.aquabot.2007.12.003
CrossRef Полный текст | Google Scholar
Loggini, B., Scartazza, A., Brugnoli, E., and Navari-Izzo, F. (1999). Система антиоксидантной защиты, пигментный состав и фотосинтетическая эффективность двух сортов пшеницы, подвергшихся засухе. Plant Physiol. 119, 1091–1099. DOI: 10.1104 / стр.119.3.1091
CrossRef Полный текст | Google Scholar
Махмуд, К., Сарвар, Г., Хуссейн, Н., Шмейский, Х., и Мухаммад, С. (2009). Влияние засоленности и содовости почвы на ростовые параметры Acacia ampiceps . Пак. J. Agric. Res. 22, 132–139.
Google Scholar
Маркар, Н. Э., и Кроуфорд, Д. Ф. (2011). Внутриспецифическая изменчивость реакции на соли и заболачивание у проростков Acacia ampiceps Maslin. Новый для. 41, 207–219. DOI: 10.1007 / s11056-010-9221-0
Новый для. 41, 207–219. DOI: 10.1007 / s11056-010-9221-0
CrossRef Полный текст | Google Scholar
Marcar, N.E., Dart, P., and Sweeney, C. (1991). Влияние засоления корневой зоны на рост и химический состав Acacia ampiceps B.R. Maslin, A. auriculiformis A. Cunn. Ex Benth. и A. mangium Willd. на двух уровнях азота. New Phytol. 119, 567–573. DOI: 10.1111 / j.1469-8137.1991.tb01049.x
CrossRef Полный текст | Google Scholar
Максвелл, К.и Джонсон Г. Н. (2000). Флуоресценция хлорофилла — практическое руководство. J. Exp. Бот. 51, 659–668. DOI: 10.1093 / jexbot / 51.345.659
CrossRef Полный текст | Google Scholar
Мимура, Т., Кура-Хотта, М., Цуджимура, Т., Охниши, М., Миура, М., Окадзаки, Ю., и др. (2003). Быстрое увеличение вакуолярного объема в ответ на солевой стресс. Planta 216, 397–402. DOI: 10.1007 / s00425-002-0878-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мораис, М. К., Пануччио М. Р., Мусколо А. и Фрейтас Х. (2012). Признаки солеустойчивости увеличивают инвазионный успех Acacia longifolia в португальских прибрежных дюнах. Plant Physiol. Biochem. 55, 60–65. DOI: 10.1016 / j.plaphy.2012.03.013
К., Пануччио М. Р., Мусколо А. и Фрейтас Х. (2012). Признаки солеустойчивости увеличивают инвазионный успех Acacia longifolia в португальских прибрежных дюнах. Plant Physiol. Biochem. 55, 60–65. DOI: 10.1016 / j.plaphy.2012.03.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Панта, С., Флауэрс, Т., Лейн, П., Дойл, Р., Харос, Г., и Шабала, С. (2014). Галофитное земледелие: истории успеха. Environ. Exp. Бот. 107, 71–83.DOI: 10.1016 / j.envexpbot.2014.05.006
CrossRef Полный текст | Google Scholar
Парида А. К., Дас А. Б. и Миттра Б. (2003). Влияние стресса NaCl на структуру, состав пигментного комплекса и фотосинтетическую активность мангровых зарослей Bruguiera parviflora . Photosynthetica 41, 191–200. DOI: 10.1023 / B: PHOT.0000011951.37231.69
CrossRef Полный текст | Google Scholar
Патель, А. Д., Ядейя, Х., и Панди, А. Н. (2010). Влияние засоления почвы на рост, водный статус и накопление питательных веществ в проростках Acacia auriculiformis (Fabaceae). J. Plant Nutr. 33, 914–932. DOI: 10.1080 / 011003669939
J. Plant Nutr. 33, 914–932. DOI: 10.1080 / 011003669939
CrossRef Полный текст | Google Scholar
Поттосин, И., и Маиз, Дж. (2002). Вакуолярный ионный транспорт высших растений в клеточном контексте. Acta Bot. Maxicana. 60, 37–77.
Google Scholar
Рехман, С., Харрия, П. Дж. К., Борн, В., и Уилкин, Дж. (2000). Взаимосвязь между ионами, энергией роста и засолением семян Acacia . Почва растений 230, 229–233.DOI: 10.1023 / A: 1004701231183
CrossRef Полный текст | Google Scholar
Роза М., Хилал М., Гонсалес Дж. А. и Прадо Ф. Э. (2009b). Влияние низких температур на активность ферментов, участвующих в разделении сахарозы и крахмала в сеянцах лебеды ( Chenopodium quinoa Willd.), Подвергшихся солевому стрессу и акклиматизированных к соли семядолям. Plant Physiol. Biochem. 47, 300–330. DOI: 10.1016 / j.plaphy.2008.12.001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Роза, М. , Prado, C., Podazza, G., Interdonato, R., González, J. A., Hilal, M., et al. (2009a). Растворимые сахара, метаболизм, сенсорный и абиотический стресс: сложная сеть в жизни растений. Завод Сигнал. Behav. 4, 388–393. DOI: 10.4161 / psb.4.5.8294
, Prado, C., Podazza, G., Interdonato, R., González, J. A., Hilal, M., et al. (2009a). Растворимые сахара, метаболизм, сенсорный и абиотический стресс: сложная сеть в жизни растений. Завод Сигнал. Behav. 4, 388–393. DOI: 10.4161 / psb.4.5.8294
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Shabala, S., Bose, J., and Hedrich, R. (2014). Солевые пузыри: какое значение они имеют? Trends Plant Sci. 19, 687–691. doi 10.1016 / j.tplants.2014.09.001
PubMed Аннотация | Google Scholar
Шабала, С.Н., Шабала, С. И., Мартыненко, А. И., Бабурина, О., Ньюман, И. А. (1998). Влияние засоления на биоэлектрическую активность, рост, накопление Na + и флуоресценцию хлорофилла листьев кукурузы: сравнительный обзор и перспективы скрининга. Aust. J. Plant Physiol. 25, 609–616. DOI: 10.1071 / PP97146
CrossRef Полный текст | Google Scholar
Ширази, М. У., Хан, М. А., Али, М., Муджтаба, С. М., Мумтаз, С. , Али, М. и др. (2006). Показатели роста и содержание питательных веществ некоторых солеустойчивых многоцелевых древесных пород, растущих в засоленной среде. Пак. J. Bot. 38, 1381–1388.
, Али, М. и др. (2006). Показатели роста и содержание питательных веществ некоторых солеустойчивых многоцелевых древесных пород, растущих в засоленной среде. Пак. J. Bot. 38, 1381–1388.
Google Scholar
Ширази, М. У., Ханзада, Б., Алам, С. М., Ансари, Р., Муджтаба, С. М., Али, М., и др. (2001). Сезонные колебания питательных веществ у двух видов Acacia , произрастающих в засоленной среде. Пак. J. Biol. Sci. 4, 514–517. DOI: 10.3923 / pjbs.2001.514.517
CrossRef Полный текст | Google Scholar
Сунарпи, Хори, Т., Мотода, Дж., Кубо, М., Янг, Х., Йода, К. и др. (2005). Повышенная солеустойчивость, опосредованная переносчиком AtHKT1, разгрузкой Na + из сосудов ксилемы в клетки паренхимы ксилемы. Plant J. 44, 928–938. DOI: 10.1111 / j.1365-313X.2005.02595.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Таджи, Т., Секи, М., Сато, М., Сакураи, Т., Кобаяши, М., Исияма, К. и др. (2004). Сравнительная геномика солеустойчивости между Arabidopsis и Arabidopsis -родственным кресс-салатом галофита с использованием микроматрицы Arabidopsis . Plant Physiol. 135, 1697–1709. DOI: 10.1104 / стр.104.039909
Сравнительная геномика солеустойчивости между Arabidopsis и Arabidopsis -родственным кресс-салатом галофита с использованием микроматрицы Arabidopsis . Plant Physiol. 135, 1697–1709. DOI: 10.1104 / стр.104.039909
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Такемура, Т., Ханагата, Н., Суугихара, К., Баба, С., Карубе, И., и Дубинский, З. (2000). Физиологические и биохимические реакции на солевой стресс в мангровых зарослях, Bruguiera gymnorrhiza . Aqua. Бот. 68, 15–28. DOI: 10.1016 / S0304-3770 (00) 00106-6
CrossRef Полный текст | Google Scholar
Tanaka, K., Ohta, K., Haddad, P. R., Fritz, J. S., Lee, K. P., Hasebe, K., et al. (1999). Мониторинг кислотных дождей в Восточной Азии с помощью портативного ионно-катионообменного хроматографического анализатора. J. Chromatog. 850, 311–317. DOI: 10.1016 / S0021-9673 (99) 00286-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Theerawitaya, C. , Samphumphuang, T., Cha-um, S., Yamada, N., and Takabe, T. (2014). Реакция проростков пальмы Nipa ( Nypa fruticans ) мангрового дерева на солевой стресс в горшечной культуре. Флора 209, 597–603. DOI: 10.1016 / j.flora.2014.08.004
, Samphumphuang, T., Cha-um, S., Yamada, N., and Takabe, T. (2014). Реакция проростков пальмы Nipa ( Nypa fruticans ) мангрового дерева на солевой стресс в горшечной культуре. Флора 209, 597–603. DOI: 10.1016 / j.flora.2014.08.004
CrossRef Полный текст | Google Scholar
Вера-Эстрелла, Р., Баркла, Б. Дж., Гарсиа-Рамирес, Л., и Пантоха, О. (2005). Солевой стресс у Thellungiella halophila активирует механизмы транспорта Na + , необходимые для устойчивости к солености. Plant Physiol. 139, 1507–1517. DOI: 10.1104 / стр.105.067850
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Верма, Д., Сингла-Парик, С. Л., Раджагопал, Д., Редди, М. К., и Сопори, С. К. (2007). Функциональная проверка новой изоформы антипортера Na + / H + из Pennisetum glaucum для повышения устойчивости риса к засолению. J. Biosci. 32, 621–628. DOI: 10.1007 / s12038-007-0061-9
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wang, J. , Li, S., Meng, Y., Ma, X., Lai, Y., Yang, K., et al. (2015). Транскриптомное профилирование реакции на солевой стресс у галофита Halogeton glomeratus . BMC Genome. 16: 169. DOI: 10.1186 / s12864-015-1373-z
, Li, S., Meng, Y., Ma, X., Lai, Y., Yang, K., et al. (2015). Транскриптомное профилирование реакции на солевой стресс у галофита Halogeton glomeratus . BMC Genome. 16: 169. DOI: 10.1186 / s12864-015-1373-z
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Wongpokkhom, N., Kheoruenromne, I., Суддхипракарн А. и Гилкс Р. Дж. (2008). Микроморфологические свойства засоленных почв Северо-Восточного Таиланда. Geoderma 144, 158–170. DOI: 10.1016 / j.geoderma.2007.10.026
CrossRef Полный текст | Google Scholar
Йокота, С. (2003). Связь между солеустойчивостью и накоплением пролина у австралийских видов Acacia . J. For. Res. 8, 89–93. DOI: 10.1007 / s103100300010
CrossRef Полный текст | Google Scholar
Чжан, Г.Х., Су, К., Ан, Л. Дж., И Ву, С. (2008). Характеристика и экспрессия вакуолярного гена-антипортера Na + / H + из галофита однодольных Aeluropus littoralis . Plant Physiol. Biochem. 46, 117–126. DOI: 10.1016 / j.plaphy.2007.10.022
Plant Physiol. Biochem. 46, 117–126. DOI: 10.1016 / j.plaphy.2007.10.022
CrossRef Полный текст | Google Scholar
Acacia longifolia (золотистая акация)
Генетика
Число хромосом, указанное для этого вида, составляет 2n = 26 (PROTA, 2015). A. longifolia не проявляет высокой фенотипической пластичности (Peperkorn et al., 2005).
Репродуктивная биология
A. longifolia дает колосовидные цветочные головки с запахом фиалки. У каждого цветка много пыльников. Как и другие австралийские акации, его опыляют самые разные универсальные насекомые, часто местные пчелы. Apis mellifera считается одним из опылителей. Хотя A. longifolia является ауткроссером и имеет низкую способность к самоопылению; в отсутствие переносчиков пыльцы произойдет некоторое самооплодотворение (Correia et al. , 2014). A. longifolia — поликарпический вид, производящий семена ежегодно на протяжении всей своей жизни. Семена эллиптические; 4-6 × 2-2,5 мм, блестящие, по 20-30 мг. Жизнеспособность семян высокая (около 90%). Средний вес 1000 семян: 16,81 г; содержание белка 13,02% (ПРОТА, 2015). Результаты тестов на всхожесть показывают 12% всхожести после 50 лет открытого хранения при комнатной температуре (PROTA, 2015).
Физиология и фенология
A. longifolia — вечнозеленое дерево, вырастающее до 9 м со средней скоростью.Кора содержит 15% дубильных веществ. Может респирать из базы. В отношении производства цветов и фруктов PROTA (2015) сообщает следующее: Цветение в Австралии: июль-ноябрь. Цветение в ЮАР: июнь-сентябрь. Плодотворение в Австралии: Лето. Плодоношение в ЮАР: ноябрь-декабрь. В Калифорнии цветение происходит с января по апрель (Baldwin et al., 2012).
Ассоциации
 org/cet»>
Этот вид способен обильно клубиться, что способствует распространению на бедных почвах (Rodríguez-Echeverría et al., 2009). В Португалии Родригес-Эчеверриа и др. (2007) сообщают, что A. longifolia ассоциируется с клубеньковыми бактериями Bradyrhizobium sp., Которые, как полагают, были занесены из Австралии с акацией.
org/cet»>
Этот вид способен обильно клубиться, что способствует распространению на бедных почвах (Rodríguez-Echeverría et al., 2009). В Португалии Родригес-Эчеверриа и др. (2007) сообщают, что A. longifolia ассоциируется с клубеньковыми бактериями Bradyrhizobium sp., Которые, как полагают, были занесены из Австралии с акацией.
Требования к окружающей среде
A. longifolia предпочитает хорошо дренированные легкие супеси и может расти на бедных питательными веществами почвах. Почвы могут быть кислыми, нейтральными или щелочными (щелочными), и растение хорошо переносит засоленные почвы.Не может расти в тени. Этот вид может противостоять сильным ветрам, но не морскому воздействию. Среднегодовая температура составляет от 10 до 19 ° C, хотя может обитать до 25 ° C. Морозостоек до -6 ° C (ПРОТА, 2015). Он устойчив к засушливым периодам, морозам и морским брызгам, но, как правило, для распространения требуется не менее 550 мм осадков в год (GISD, 2015).
Большинство семян имеют врожденный покой и нуждаются в стимуляции, чтобы вызвать прорастание, обычно огнем (Dennil et al, 1993; Marchante et al ., 2010 ) . Тепло увеличивает всхожесть семян A. longifolia , которые могут выдерживать максимальную температуру до 160 ° C более 20 минут (Behenna et al., 2008).
. зеленые листья, зеленые годы, зеленые фрукты, зеленый учитель
. наречия
. предлоги
. местоимения
2. Составляющие предикативной фразы
. не может быть изменено
.характеризуются интонацией
. изменять друг друга
. равны в ранге
3. Выбрать набор слов, в которых соблюдаются синтагматические отношения?
. сумка, портфель, сумочка, кошелек, рюкзак, чемодан
. зеленые листья, зеленые годы, зеленые плоды, зеленый учитель
. привет, привет, до свидания, до скорых встреч, все в порядке
. учитель, коуч, лектор, инструктор, тьютор, педагог
учитель, коуч, лектор, инструктор, тьютор, педагог
4.Как составлены подчеркнутые слова: милое лицо, дружеский визит?
. компаундирование
. преобразование
. обратное формирование
. суффикс
5. Каково происхождение аффиксов ism, -ics, -ist, poly-, dis-?
. Латинский
. Французы
. Скандинавский
. Греческий
6. Структурно морфемы делятся на
..продуктивно и непродуктивно
. свободный, переплетенный, полусвязанный
. словообразовательные и флексионные
. корни и аффиксы
7. Какие морфы характеризуются следующим определением: «Они идентичны по значению и имеют разные фонетические формы в разных контекстах»?
. аллофоны
. дополнительные морфы
. корневые морфы
. алломорфы
8.Как происходит процесс образования омонимов каб (кабриолет) — капуста (капуста)
. раскол полисемии
. укорочение
. заимствование
. преобразование
9. Какая линия отвечает за метонимию?
. Я никогда не читал Бальзака в оригинале
. У него была яйцеобразная голова и лягушачьи челюсти
. Какая буря аплодисментов!
.Мое сердце бьется от волнения.
10. В какой строке находятся лексические омонимы (одна и та же часть речи)?
. брошенный на трон, пила, голый медведь
. гвоздь — гвоздь, банка — банка, двор — двор
. спросила — спросила, поставила — поставила, братья
. какая — сумма, так — пила, грипп — прилетела
11. Что такое «синонимическая доминанта»?
. общий термин, нейтральный по стилю и с большой комбинационной способностью
.структурно непродуктивное слово
. это заемное слово
. этимологически настоящее слово
12. Отличие фразеологизмов от свободных словосочетаний заключается в
Отличие фразеологизмов от свободных словосочетаний заключается в
. синтаксические особенности (невозможность преобразований)
. семантические особенности (частично или полностью немотивированные)
. морфологические особенности
. синтаксические и семантические особенности?
13.Можно использовать архаизмы
. для создания исторической атмосферы
. показать, что говорящий привязан к употреблению необычных слов
. чтобы избежать тавтологии
. произвести юмористический эффект
14. Какие из слов родные?
. курица, корова, коза, ворона, птица, медведь, лиса, заяц, жаворонок
. голубь, индейка, кенгуру, жираф, белка, зебра
.пришел, крокодил, гиена, горилла, рысь, обезьяна
. картофель, помидор, табак, мачо, фиеста
15. Найдите правильный тип преобразования слов, приведенных ниже: круговой, преступник преступник, сказать, сказать, попытаться
. вербализация
вербализация
. обоснование
. адъективизация
. реклама
16. Какие слова образуют образцы слов: грабить, редактировать, кататься на коньках, благожелать, вызывать энтузиазм?
.преобразование
. бэк-формирование
. проставление
. Редупликация
17. Какая из перечисленных ниже групп слов соответствует пассивному словарю?
. архаизмы, диалектные слова, заимствования
. заимствования и сленговые слова
. профессионализм, варварство, заимствования
. неологизмы, историзмы, архаизмы
18.Коннотативный компонент — это то, что предлагается или связано с
.. определенное слово, означающее
. определенная концепция
. определенный символ
. некий референт
19. Какая линия составляет омографы?
. кроватка — детская кроватка, игра — игра, матч матч
. полюс — опрос, аромат — отправлено, равнина — самолет
полюс — опрос, аромат — отправлено, равнина — самолет
. брошенный на трон, пила, голый медведь
.бас — окунь, пустыня — пустыня, фуршет — буфет
20. Синонимы, принадлежащие к одному стилистическому слою, имеющие одинаковую коннотацию, которые характеризуются различием в дифференцирующих семах денотационного компонента их лексического значения, именуются
. стилистический
. абсолютный
. идеографический
. фразеологический
21. Какие виды фразеологизмов делали акад.Выделяете В.Виноградова?
. фраземы и идиомы
. именительный и коммуникативный
. фразеологические сочетания, фразеологические единства, фразеологические сочетания
. одно- и двухвершинные блоки
22. Какие системные отношения основаны на линейности речи?
. синтагматический
. синоним
.деривационная
. парадигматический
парадигматический
23. Слова, которые встречаются в нескольких языках в результате заимствования из одного и того же источника, — это
. международные слова
. историзмы
. архаичные слова
. новые слова
24. Какая группа слов относится к исконному стоку?
. слива, пальма, акация, сосна, баобаб, мальва, груша
.картофель, помидор, табак, мачо, фиеста
. орех, желудь, пихта, грецкий орех, фундук, ясень, дуб
. абрикос, апельсин, банан, гранат, дыня, вишня, лимон
25. Под переводной ссудой понимается
. два или более слова одного и того же языка, пришедшие разными путями из одного и того же основного исходного слова
. развитие в английском слове нового значения под влиянием коррелированной единицы в другом языке
. слово или фраза, образованные из материала, доступного на данном языке, но по чужеродному образцу посредством дословного перевода каждого компонента морфема за морфемой
. слово, которое встречается в нескольких языках в результате заимствования из одного и того же основного источника
слово, которое встречается в нескольких языках в результате заимствования из одного и того же основного источника
26. Морфемы -ness, -less, -dis выделены как
.. бесплатно
. архаичный
. связанный
.полусвязанный
27. Какие морфемы выделяются семантически?
. свободный, переплетенный, полусвязанный
. корни и аффиксы
. грамматические флексии
. корни, аффиксы, флексии
28. Суффикс т.е. в тете называется
.. архаичный
. увеличивающий
. уменьшительное
. продуктивная
29.Слово — это единство звуковой формы и
. символ
. понятие
. референт
. значение
30. Раздел лексикологии, изучающий проблему лексического значения, называется
.. семасиология
. этимология
. морфология
. топонимия
топонимия
31. Какая линия соответствует метафоре?
.Яма громко аплодировали
. Он говорит как книга
. Киев — сердце нашей страны
. Кофейник кипящий
32. Какой процесс семантических изменений определяется как «процесс, в результате которого слово по той или иной причине приобретает дурную славу и становится менее респектабельным»?
. сужение значения
. передача смысла
. расширение значения
.деградация смысла
33. Какое из определений соответствует понятию «синонимы»?
. слова, для которых характерна идиоматичность
. слова, частично совпадающие по звуковой форме, но различающиеся по значению
. слова, принадлежащие к одной и той же части речи, различающиеся звуковой формой, но идентичные или близкие по значению
. слова, одинаковые по звуковой форме, но разные по значению
34. Описательная лексикология
Описательная лексикология
. структура словарных единиц
. словарный запас данного языка на данном этапе его развития
. описание характерных особенностей лексики данного языка
. история словарного запаса языка, показывающая его изменение и развитие с течением времени
35. Выберите набор слов, в которых наблюдаются парадигматические отношения
.зеленые листья, зеленые годы, зеленый человек, в длинном зеленом
. тяжелое небо, тяжелая сумка, сильный дождь, тяжелое сердце, тяжелый ужин
. рука, удобный, почерк, рукописный, горстка, гандбол ОБЩИЙ КОРЕНЬ
. белый свет, белая ворона, белая ночь, белая ложь, белый человек
36. Заимствованные слова, которые не усвоены в принимающем языке, — это
. жаргонизмы
. международный мир
.варварства
. историзмы
37. В чем разница между составными словами и именными словосочетаниями?
. в сочетании слов каждый элемент подчеркнут и записан отдельно
. у них разное значение
. они могут иметь разные части речи
. они имеют разные значения, где первый элемент изменяет другой
38. Конверсия как морфологический способ образования новых слов была предложена
..Гл. Картофель фри
. Отто Йесперсен
. А. Смирницкий
. О. Ахманова
39. Составное словообразование, где
. глаголы образуются от существительных путем сдвига основы
. слова состоят как минимум из двух основ, которые встречаются в языке как свободные формы
. новые слова образуются путем добавления аффиксов к разным основам
.новые слова образуются без добавления словообразовательных элементов
40. Термин «конверсия» впервые был введен
.. S.Ullmann
. Отто Йесперсен
. J. Lyons
. Х. Свит
41. Выберите историзмы из групп ниже
.. тореадор, раджа, шах, мэр
. мама, Ева, твоя, ты, да, нет, луна, часто
. голубь, индейка, кенгуру, жираф, белка, зебра
.анарх, барон, мушкетер, вассал, карбонари
42. Какой термин можно определить как «слова, идентичные по звуковой форме, но не имеющие общих сем или ассоциаций»
. лексические омонимы
. паронимы
. многозначные слова
. фразеологизмы
43. Как происходит процесс образования синонимов сияющий яркий?
. омонимы
.сдвиг смысла
. преобразование
. заем
44. Укажите фразеологизмы, которые считаются синонимами
. на всю толщину, всеми правдами и неправдами, ради любви или денег
. рог изобилия, Авгиевы конюшни, Троянский конь, суета тщеславия
. взять быка за рога по всем параметрам одним прыжком
.понемногу, на месте, начинать не с того конца
45. Специальная лексикология
. словарный запас данного языка на данном этапе его развития
. характерные особенности слов в лексике каждого языка
. история словарного запаса языка, показывающая его изменение и развитие с течением времени
. Испанский и португальский
.Итальянский
. Латинский
. Французы
47. Под переводной ссудой понимается
. два или более слова одного и того же языка, пришедшие разными путями из одного и того же основного исходного слова
. слово, которое встречается в нескольких языках в результате заимствования из одного и того же основного источника
. семантическое ядро
. референт
51. Денотативный компонент лексического значения
..компонент значения слова, повторяющийся в идентичных наборах грамматических форм разных слов
. лексическое ядро слова, которое связано с референтом и понятием, которое обозначает данное слово
. материальное значение слова, которое непосредственно связано с предметом или понятием, которое выражает данное слово
. минимальная отличительная единица
52. Что понимается под «эмоциональным зарядом»?
.потенциальная способность слов встречаться с другими словами
. отношение говорящего к тому, о чем говорят
. оттенки значения или разные степени заданного качества
. социальная сфера, в которой происходит дискурс
53. Что такое словоформа?
. систематическое формо-смысловое соответствие между словами
. значимая часть слова
.фонемы
. морфемы
56. «Сломать — сломаться» представляет
. постепенное противодействие
. приватная оппозиция
. морфологический принцип
. синтаксические позиции слов
. синтаксический принцип
. морфологические и функциональные принципы
59. Категория аспекта представлена противопоставлением
.общие и непрерывные аспекты
. прямой и наклонный аспекты
. неопределенные и совершенные аспекты
. Совершенные и несовершенные аспекты
60. Инфинитив имеет такие грамматические категории, как
.. время и вид
. по согласованию
62. Словосочетание
.. лишен именительной функции
.коммуникативная единица языка
. именная единица
. речевой акт
63. Ch. Фрайз отличает отнесение слов к определенной группе.
. 4 класса
. 6 классов
. 3 класса
. 5 классов
64. Традиционно части речи рассматриваются как
.. функциональные классы
. синтаксические классы
.корпус и номер
. пол и корпус
. время и вид
. число и настроение
67. Проф. Ильиш выделяет
. четыре формы вида времени: неопределенное, непрерывное, совершенное, совершенное непрерывное
. голос и корреляция
70. Какое из явлений не является характерной чертой словосочетания?
. договор
.предсказуемость
. подчинение
. модальность
71. Прогнозируемость
. отношение мысли, выраженной в предложении, к ситуации речи
. связь между названными объектами и реальностью
. отношение говорящего к тому, о чем говорится
. фонематически обусловлен
. заменяющий
.морфологически обусловлено
. непроизводственные элементы
73. Непрерывные морфемы по М.Блоху —
.. аналитические флексии
. вспомогательное оборудование
. аффиксы
. дополнительная
74. У абстрактных существительных много общего с
.. исчисляемые существительные
. имена собственные
. конкретные существительные
.несчетные существительные
75. Какая теория утверждает, что падеж зависит от положения существительного в предложении?
. лексико-функциональный
77. Прилагательное может выполнять синтаксическую функцию
.. наречный модификатор и атрибут
. объект и предикатив
. подлежащее и сказуемое
. наречия
.предлоги
. местоимения
2. Составляющие предикативной фразы
. не может быть изменено
. характеризуются интонацией
. изменять друг друга
. равны в ранге
3. Выбрать набор слов, в которых соблюдаются синтагматические отношения?
. сумка, портфель, сумочка, кошелек, рюкзак, чемодан
.зеленые листья, зеленые годы, зеленые плоды, зеленый учитель
. привет, привет, до свидания, до скорых встреч, все в порядке
. учитель, коуч, лектор, инструктор, тьютор, педагог
4. Как составлены подчеркнутые слова: милое лицо, дружеский визит?
. компаундирование
. преобразование
. обратное формирование
. суффикс
5. Каково происхождение аффиксов ism, -ics, -ist, poly-, dis-?
.Латинский
. Французы
. Скандинавский
. Греческий
6. Структурно морфемы делятся на
.. продуктивно и непродуктивно
. свободный, переплетенный, полусвязанный
. словообразовательные и флексионные
. корни и аффиксы
7. Какие морфы характеризуются следующим определением: «Они идентичны по значению и имеют разные фонетические формы в разных контекстах»?
.